

Emergence of multiplicative auditory responses in the midbrain of the barn owl
- PMID: 17615132
- PMCID: PMC2532518
- DOI: 10.1152/jn.00370.2007
Emergence of multiplicative auditory responses in the midbrain of the barn owl
Abstract
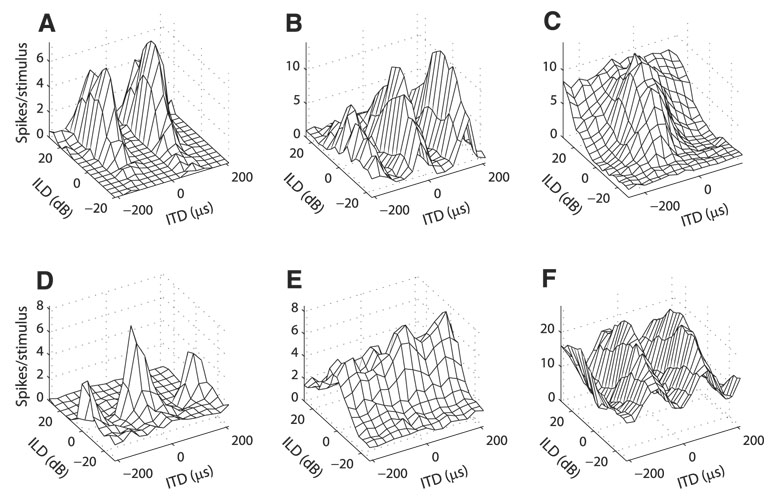
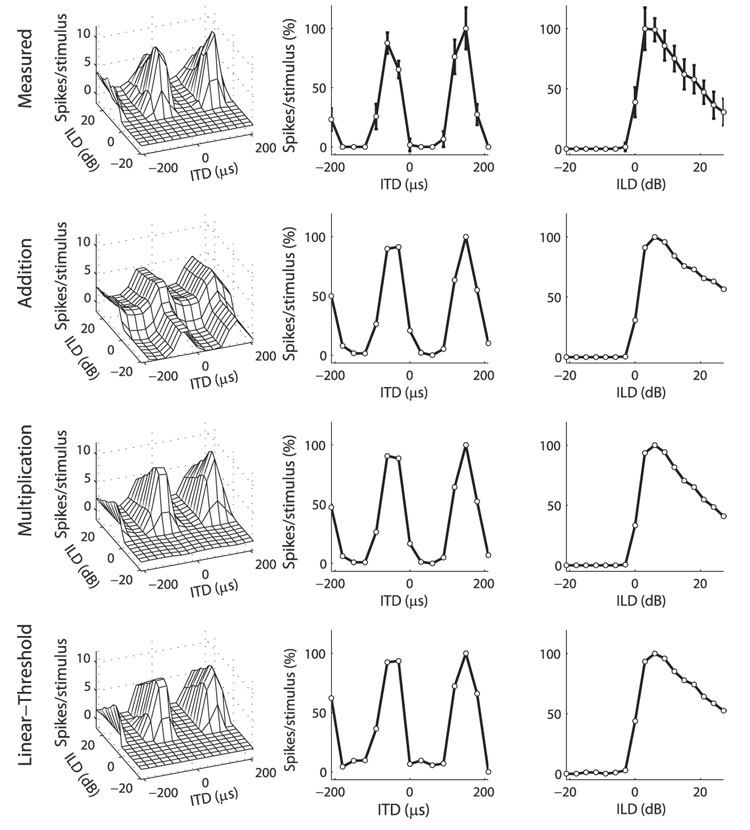
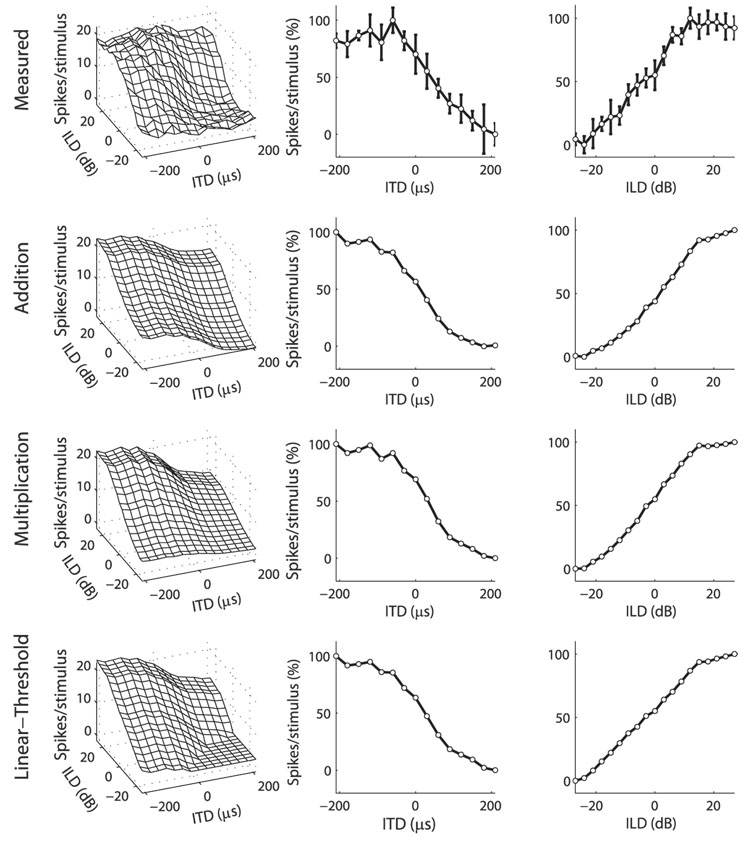
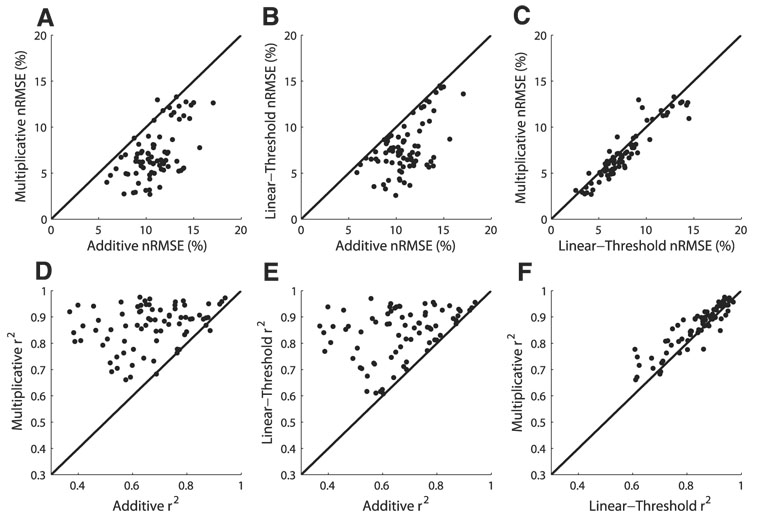
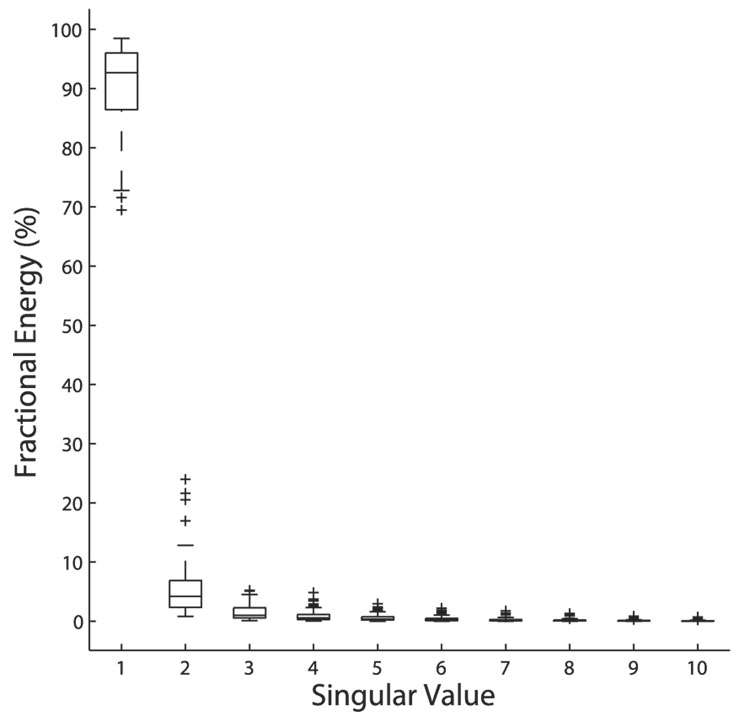
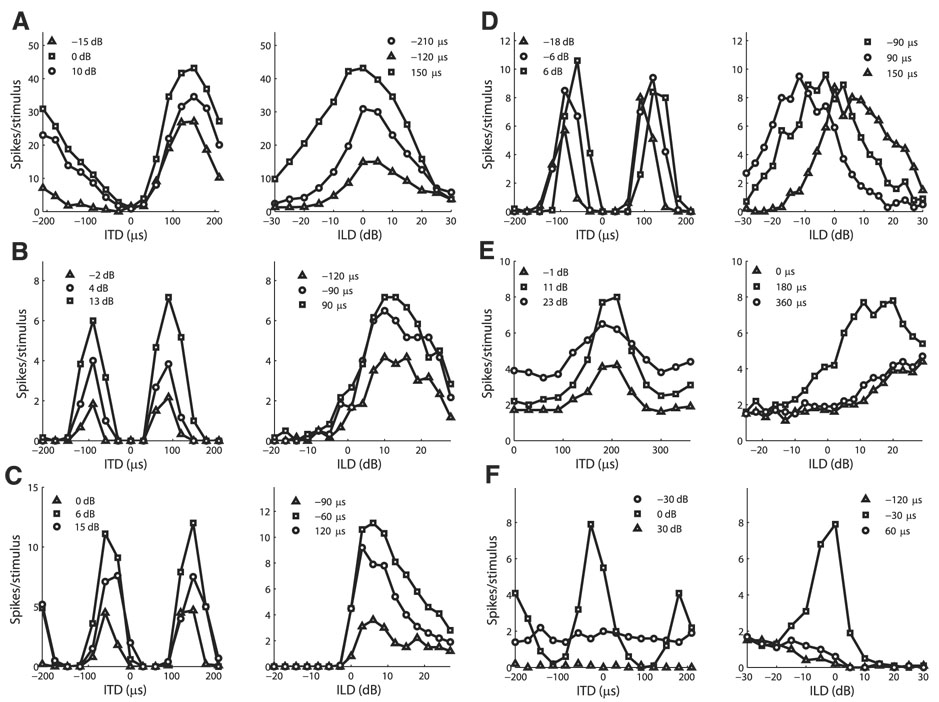
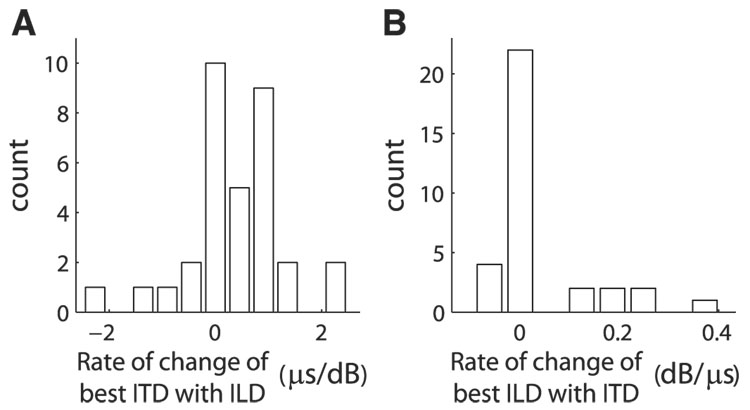
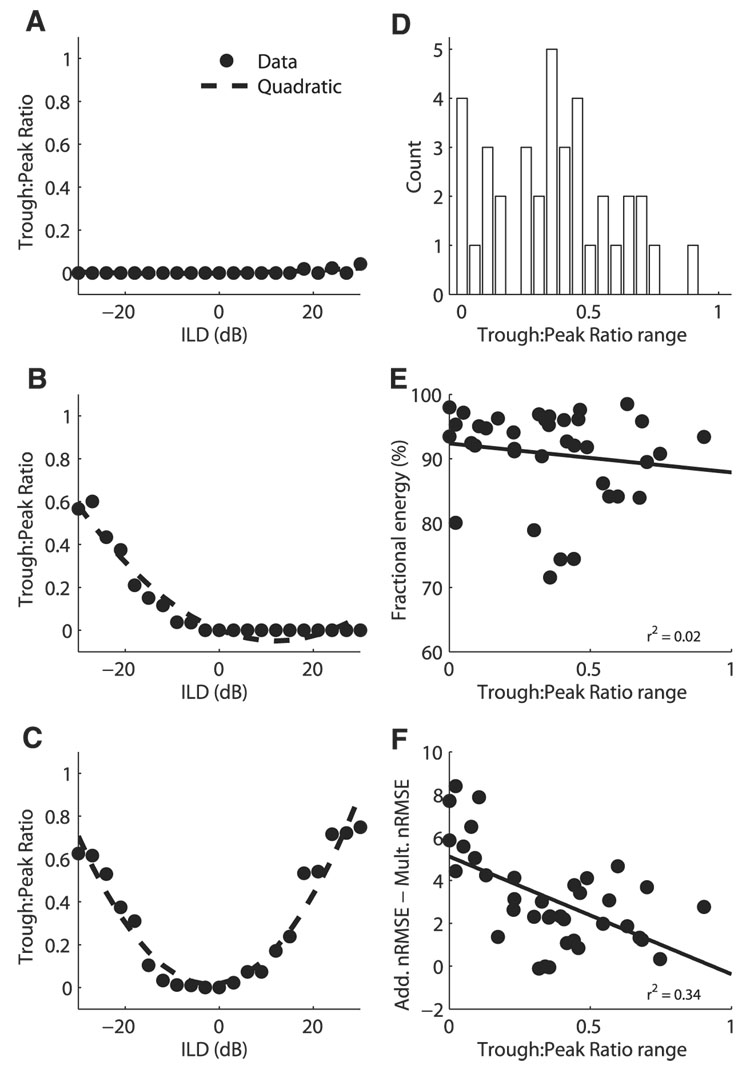
Space-specific neurons in the barn owl's auditory space map gain spatial selectivity through tuning to combinations of the interaural time difference (ITD) and interaural level difference (ILD). The combination of ITD and ILD in the subthreshold responses of space-specific neurons in the external nucleus of the inferior colliculus (ICx) is well described by a multiplication of ITD- and ILD-dependent components. It is unknown, however, how ITD and ILD are combined at the site of ITD and ILD convergence in the lateral shell of the central nucleus of the inferior colliculus (ICcl) and therefore whether ICx is the first site in the auditory pathway where multiplicative tuning to ITD- and ILD-dependent signals occurs. We used extracellular recording of single neurons to determine how ITD and ILD are combined in ICcl of the anesthetized barn owl (Tyto alba). A comparison of additive, multiplicative, and linear-threshold models of neural responses shows that ITD and ILD are combined nonlinearly in ICcl, but the interaction of ITD and ILD is not uniformly multiplicative over the sample. A subset (61%) of the neural responses is well described by the multiplicative model, indicating that ICcl is the first site where multiplicative tuning to ITD- and ILD-dependent signals occurs. ICx, however, is the first site where multiplicative tuning is observed consistently. A network model shows that a linear combination of ICcl responses to ITD-ILD pairs is sufficient to produce the multiplicative subthreshold responses to ITD and ILD seen in ICx.
Figures
Similar articles
-
Multiplicative auditory spatial receptive fields created by a hierarchy of population codes.PLoS One. 2009 Nov 24;4(11):e8015. doi: 10.1371/journal.pone.0008015. PLoS One. 2009. PMID: 19956693 Free PMC article.
-
Comparison of midbrain and thalamic space-specific neurons in barn owls.J Neurophysiol. 2006 Feb;95(2):783-90. doi: 10.1152/jn.00833.2005. J Neurophysiol. 2006. PMID: 16424454 Free PMC article.
-
Auditory spatial tuning at the crossroads of the midbrain and forebrain.J Neurophysiol. 2009 Sep;102(3):1472-82. doi: 10.1152/jn.00400.2009. Epub 2009 Jul 1. J Neurophysiol. 2009. PMID: 19571193 Free PMC article.
-
Binaural processing in the synthesis of auditory spatial receptive fields.Biol Cybern. 2003 Nov;89(5):371-7. doi: 10.1007/s00422-003-0442-6. Epub 2003 Nov 4. Biol Cybern. 2003. PMID: 14669017 Review.
-
The synthesis and use of the owl's auditory space map.Biol Cybern. 2003 Nov;89(5):378-87. doi: 10.1007/s00422-003-0443-5. Epub 2003 Dec 4. Biol Cybern. 2003. PMID: 14669018 Review.
Cited by
-
Optimal nonlinear cue integration for sound localization.J Comput Neurosci. 2017 Feb;42(1):37-52. doi: 10.1007/s10827-016-0626-4. Epub 2016 Oct 6. J Comput Neurosci. 2017. PMID: 27714569 Free PMC article.
-
Two Types of Auditory Spatial Receptive Fields in Different Parts of the Chicken's Midbrain.J Neurosci. 2022 Jun 8;42(23):4669-4680. doi: 10.1523/JNEUROSCI.2204-21.2022. Epub 2022 May 4. J Neurosci. 2022. PMID: 35508384 Free PMC article.
-
A Physiologically Inspired Model for Solving the Cocktail Party Problem.J Assoc Res Otolaryngol. 2019 Dec;20(6):579-593. doi: 10.1007/s10162-019-00732-4. Epub 2019 Aug 7. J Assoc Res Otolaryngol. 2019. PMID: 31392449 Free PMC article.
-
Multiplicative auditory spatial receptive fields created by a hierarchy of population codes.PLoS One. 2009 Nov 24;4(11):e8015. doi: 10.1371/journal.pone.0008015. PLoS One. 2009. PMID: 19956693 Free PMC article.
-
The representation of sound localization cues in the barn owl's inferior colliculus.Front Neural Circuits. 2012 Jul 11;6:45. doi: 10.3389/fncir.2012.00045. eCollection 2012. Front Neural Circuits. 2012. PMID: 22798945 Free PMC article.
References
-
- Andersen RA, Snyder LH, Bradley DC, Xing J. Multimodal representation of space in the posterior parietal cortex and its use in planning movements. Annu Rev Neurosci. 1997;20:303–330. - PubMed
-
- Arthur BJ. Sensitivity to spectral interaural intensity difference cues in space-specific neurons of the barn owl. J Comp Physiol A Sens Neural Behav Physiol. 2004;190:91–104. - PubMed
-
- Egelhaaf M, Borst A, Reichardt W. Computational structure of a biological motion-detection system as revealed by local detector analysis in the fly’s nervous system. J Opt Soc Am. 1989;6:1070–1087. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous