

Inhibiting endoplasmic reticulum (ER)-associated degradation of misfolded Yor1p does not permit ER export despite the presence of a diacidic sorting signal
- PMID: 17615300
- PMCID: PMC1951769
- DOI: 10.1091/mbc.e07-01-0046
Inhibiting endoplasmic reticulum (ER)-associated degradation of misfolded Yor1p does not permit ER export despite the presence of a diacidic sorting signal
Abstract
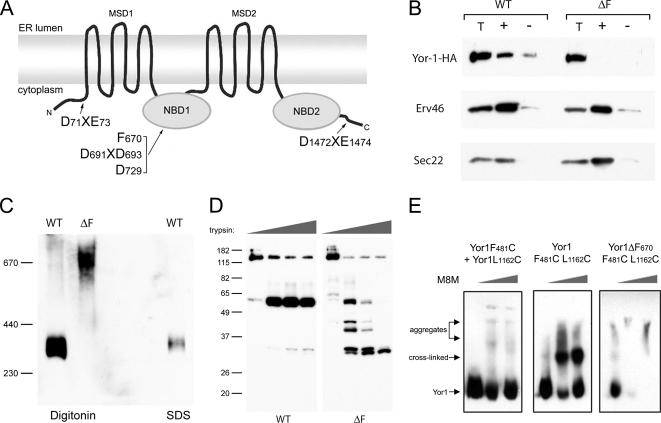
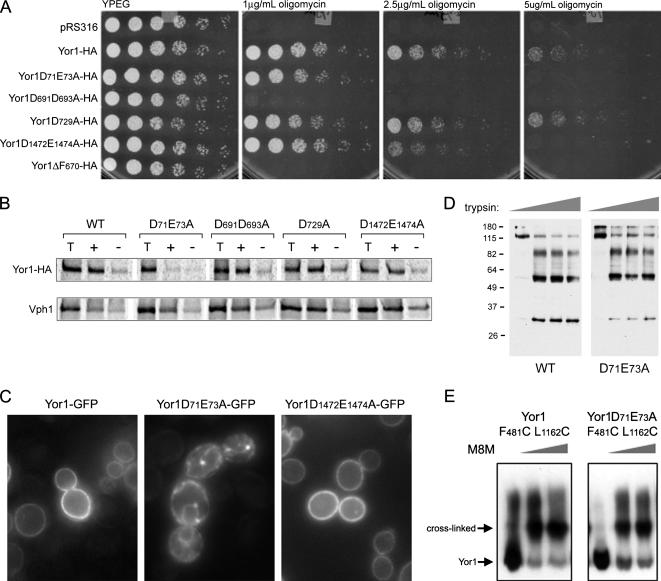
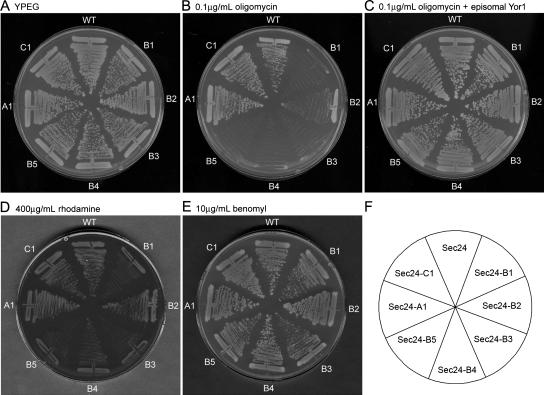
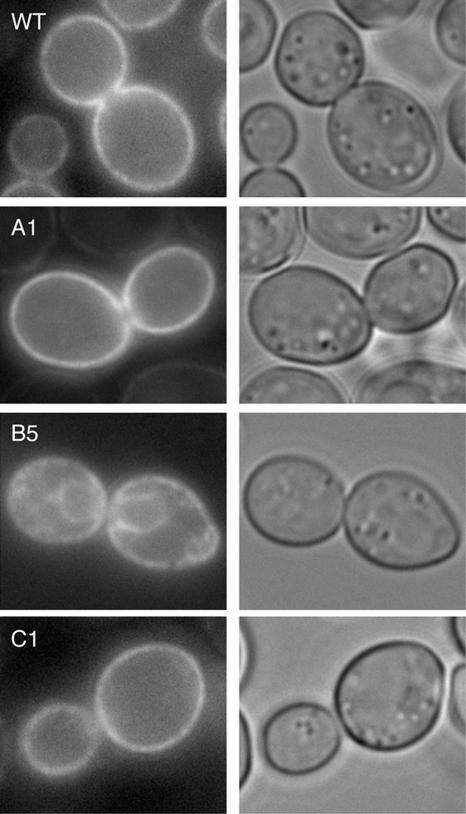
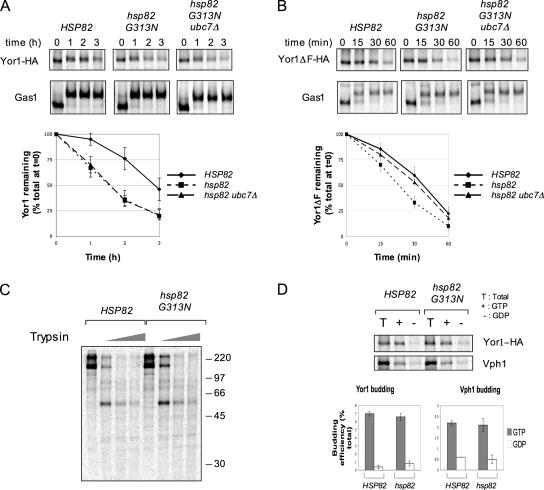
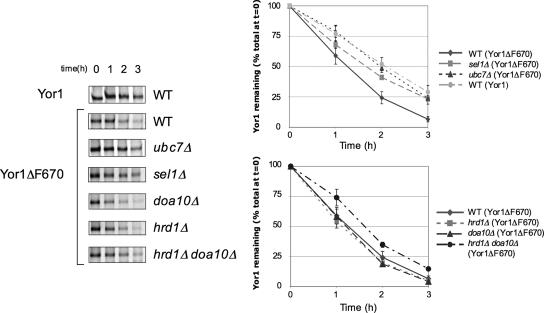
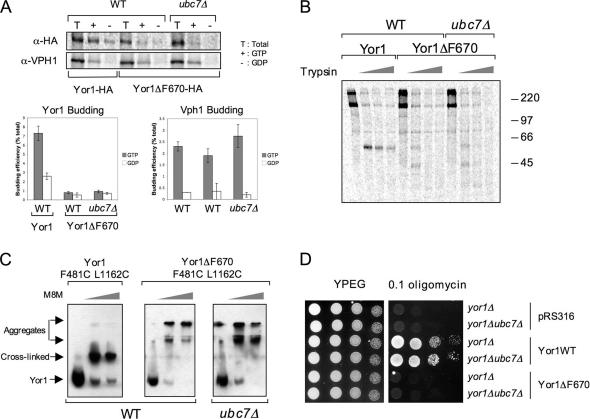
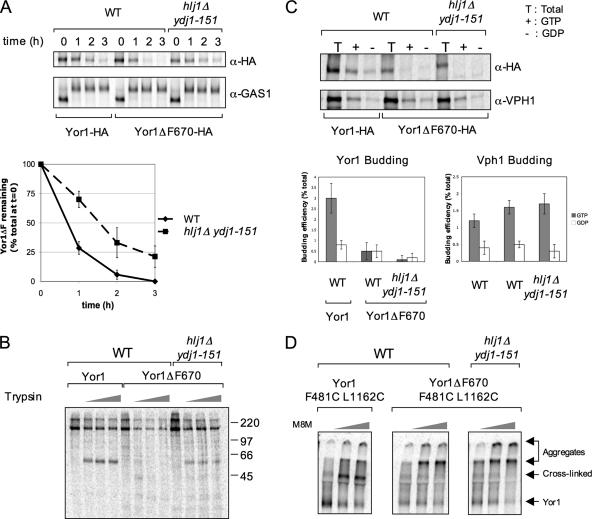
Capture of newly synthesized proteins into endoplasmic reticulum (ER)-derived coat protomer type II (COPII) vesicles represents a critical juncture in the quality control of protein biogenesis within the secretory pathway. The yeast ATP-binding cassette transporter Yor1p is a pleiotropic drug pump that shows homology to the human cystic fibrosis transmembrane conductance regulator (CFTR). Deletion of a phenylalanine residue in Yor1p, equivalent to the major disease-causing mutation in CFTR, causes ER retention and degradation via ER-associated degradation. We have examined the relationship between protein folding, ERAD and forward transport during Yor1p biogenesis. Uptake of Yor1p into COPII vesicles is mediated by an N-terminal diacidic signal that likely interacts with the "B-site" cargo-recognition domain on the COPII subunit, Sec24p. Yor1p-DeltaF is subjected to complex ER quality control involving multiple cytoplasmic chaperones and degradative pathways. Stabilization of Yor1p-DeltaF by inhibiting its degradation does not permit access of Yor1p-DeltaF to COPII vesicles. We propose that the ER quality control checkpoint engages misfolded Yor1p even after it has been stabilized by inhibition of the degradative pathway.
Figures
Similar articles
-
Traffic-independent function of the Sar1p/COPII machinery in proteasomal sorting of the cystic fibrosis transmembrane conductance regulator.J Cell Biol. 2003 Jan 20;160(2):157-63. doi: 10.1083/jcb.200210086. Epub 2003 Jan 21. J Cell Biol. 2003. PMID: 12538638 Free PMC article.
-
Substitution of Yor1p NBD1 residues improves the thermal stability of Human Cystic Fibrosis Transmembrane Conductance Regulator.Protein Eng Des Sel. 2017 Oct 1;30(10):729-741. doi: 10.1093/protein/gzx054. Protein Eng Des Sel. 2017. PMID: 29053845 Free PMC article.
-
Intragenic suppressing mutations correct the folding and intracellular traffic of misfolded mutants of Yor1p, a eukaryotic drug transporter.J Biol Chem. 2010 Nov 19;285(47):36304-14. doi: 10.1074/jbc.M110.142760. Epub 2010 Sep 13. J Biol Chem. 2010. PMID: 20837481 Free PMC article.
-
Mechanisms of COPII vesicle formation and protein sorting.FEBS Lett. 2007 May 22;581(11):2076-82. doi: 10.1016/j.febslet.2007.01.091. Epub 2007 Feb 14. FEBS Lett. 2007. PMID: 17316621 Review.
-
ER-associated degradation of membrane proteins in yeast.ScientificWorldJournal. 2006 Aug 17;6:967-83. doi: 10.1100/tsw.2006.191. ScientificWorldJournal. 2006. PMID: 16921443 Free PMC article. Review.
Cited by
-
Cadmium-mediated rescue from ER-associated degradation induces expression of its exporter.Proc Natl Acad Sci U S A. 2009 Jun 23;106(25):10189-94. doi: 10.1073/pnas.0812114106. Epub 2009 Jun 10. Proc Natl Acad Sci U S A. 2009. PMID: 19515821 Free PMC article.
-
Recent Insights into the Role of Unfolded Protein Response in ER Stress in Health and Disease.Front Cell Dev Biol. 2017 May 10;5:48. doi: 10.3389/fcell.2017.00048. eCollection 2017. Front Cell Dev Biol. 2017. PMID: 28540288 Free PMC article. Review.
-
Secretory protein biogenesis and traffic in the early secretory pathway.Genetics. 2013 Feb;193(2):383-410. doi: 10.1534/genetics.112.142810. Genetics. 2013. PMID: 23396477 Free PMC article. Review.
-
A yeast phenomic model for the gene interaction network modulating CFTR-ΔF508 protein biogenesis.Genome Med. 2012 Dec 27;4(12):103. doi: 10.1186/gm404. Genome Med. 2012. PMID: 23270647 Free PMC article.
-
Targeted degradation of ABC transporters in health and disease.J Bioenerg Biomembr. 2007 Dec;39(5-6):489-97. doi: 10.1007/s10863-007-9120-z. J Bioenerg Biomembr. 2007. PMID: 17972020 Review.
References
-
- Ahner A., Brodsky J. L. Checkpoints in ER-associated degradation: excuse me, which way to the proteasome? Trends Cell Biol. 2004;14:474–478. - PubMed
-
- Barlowe C., Orci L., Yeung T., Hosobuchi M., Hamamoto S., Salama N., Rexach M. F., Ravazzola M., Amherdt M., Schekman R. COPII: a membrane coat formed by Sec proteins that drive vesicle budding from the endoplasmic reticulum. Cell. 1994;77:895–907. - PubMed
-
- Bukau B., Weissman J., Horwich A. Molecular chaperones and protein quality control. Cell. 2006;125:443–451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
