

Deamidation alters the structure and decreases the stability of human lens betaA3-crystallin
- PMID: 17616172
- PMCID: PMC2597435
- DOI: 10.1021/bi700487q
Deamidation alters the structure and decreases the stability of human lens betaA3-crystallin
Abstract
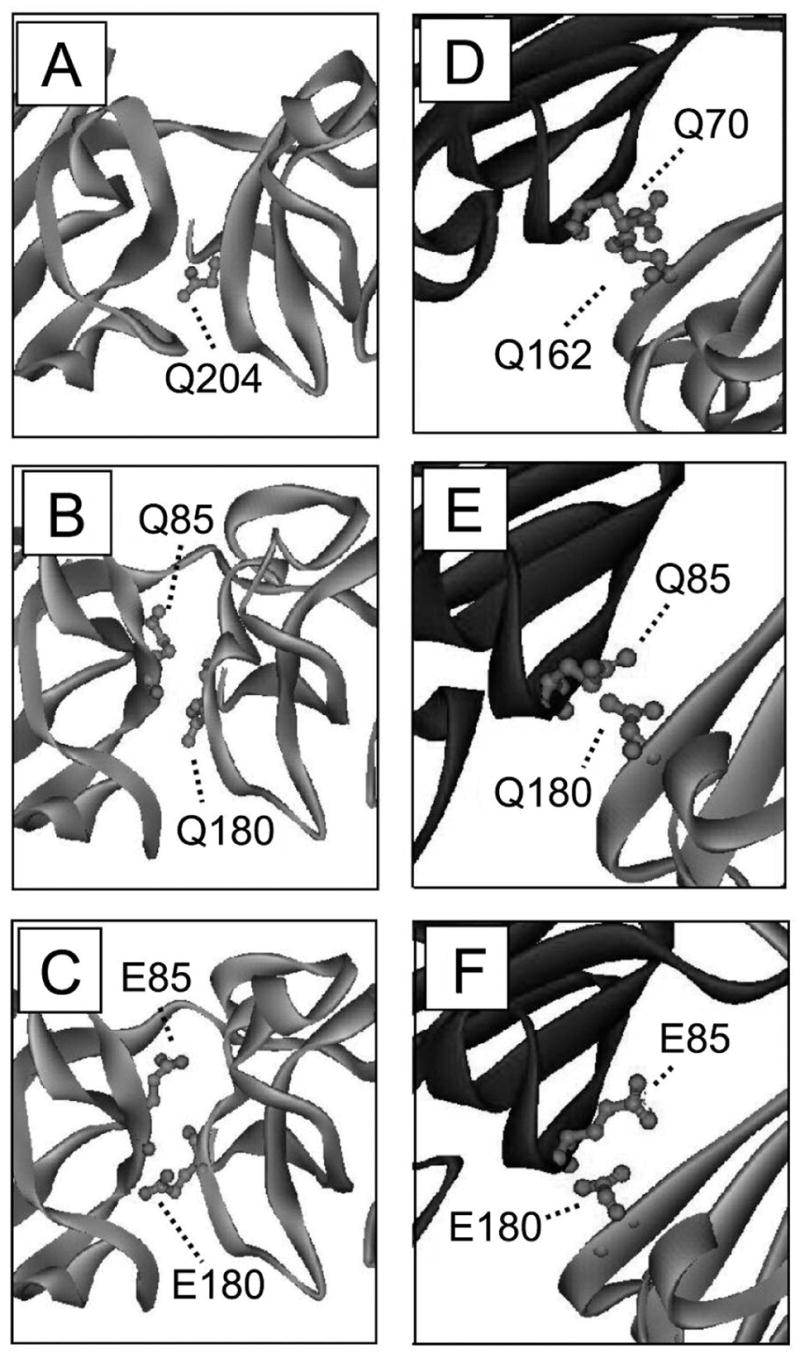
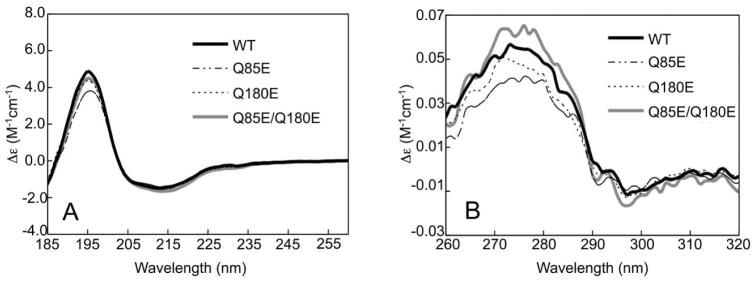
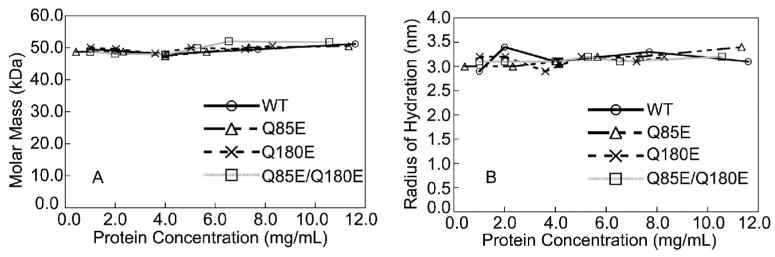
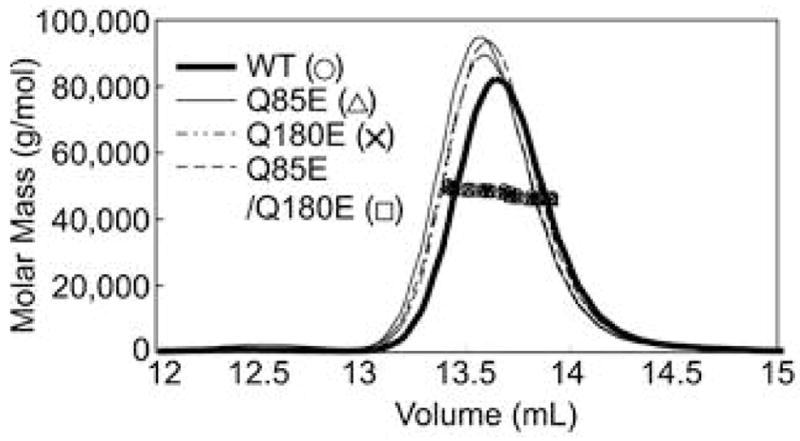
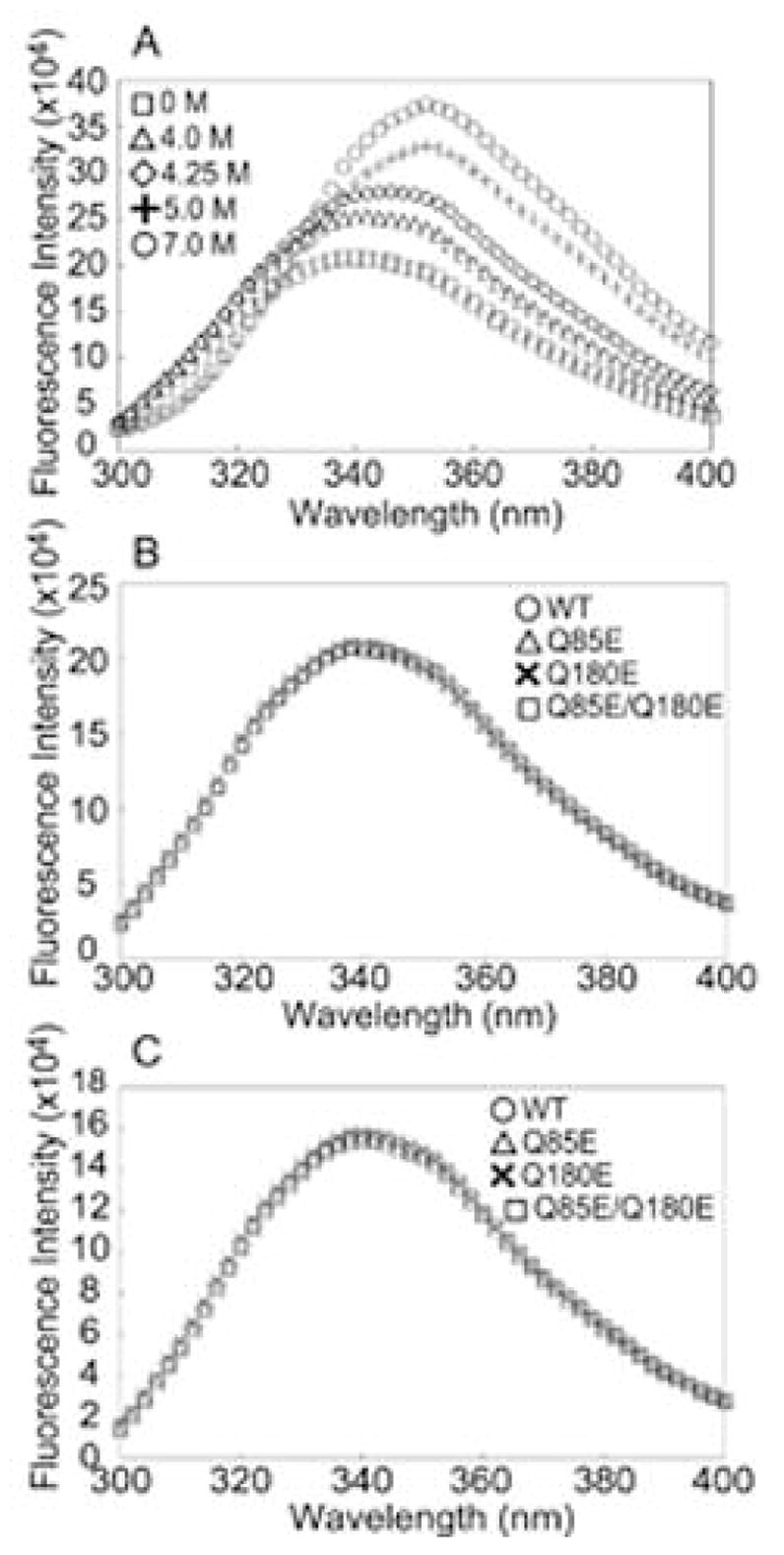
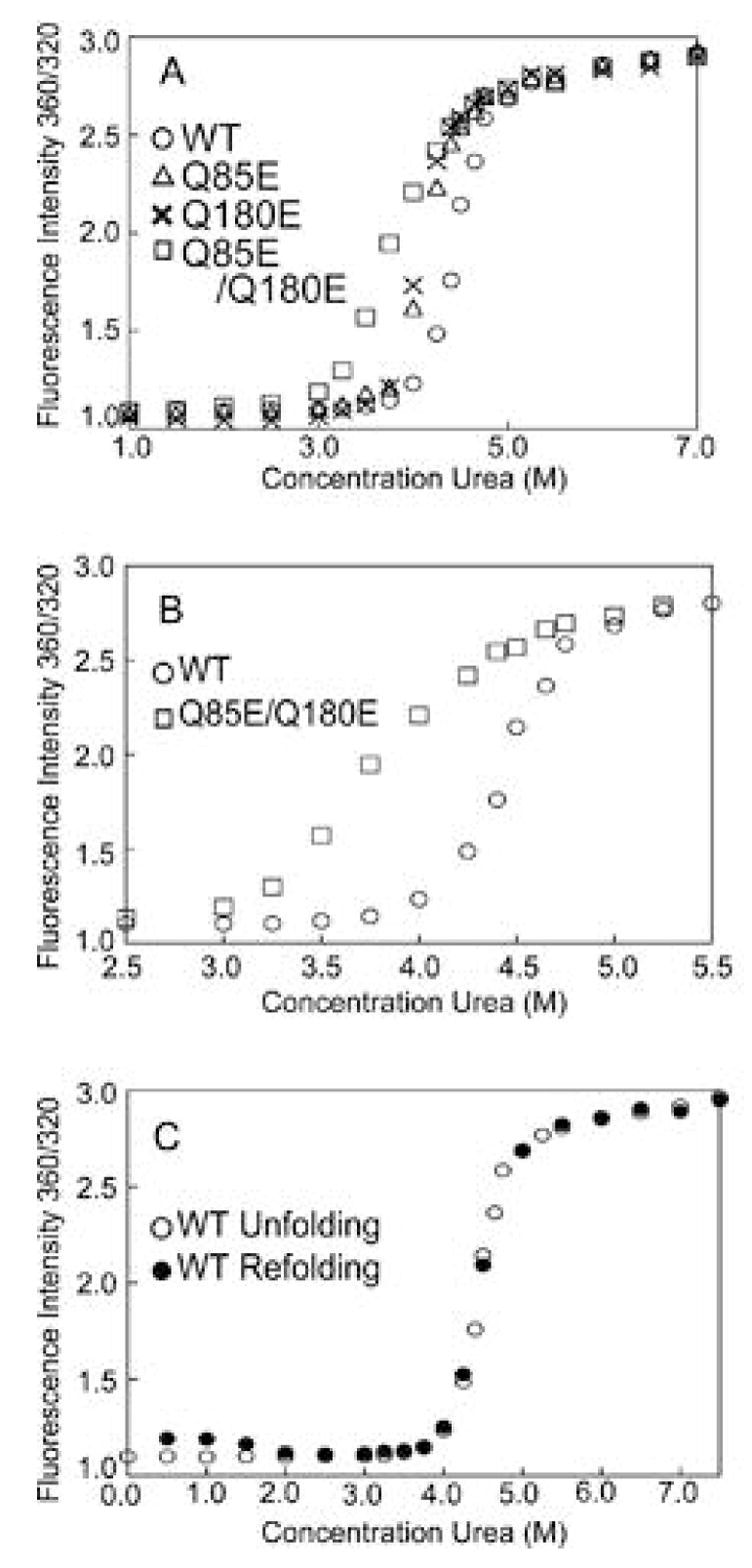
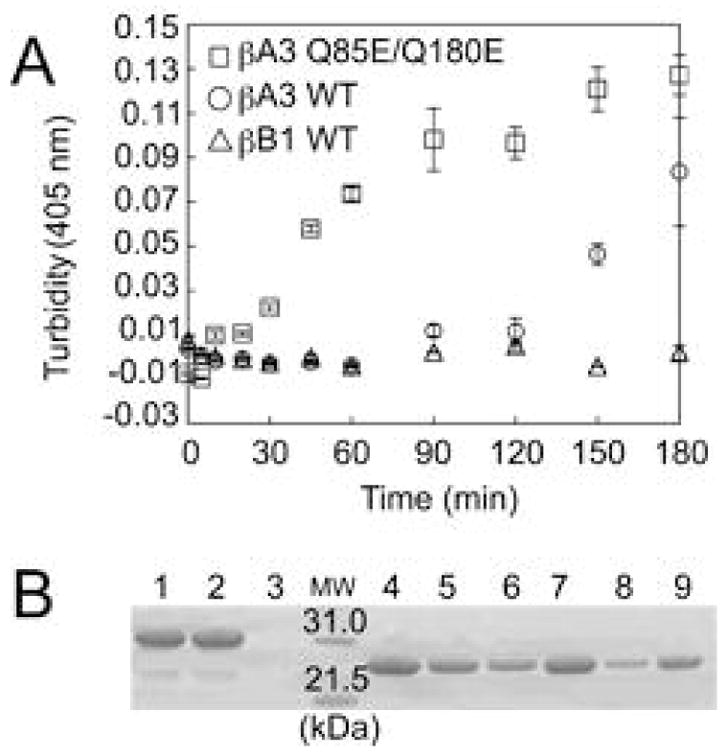
According to the World Health Organization, cataracts account for half of the blindness in the world, with the majority occurring in developing countries. A cataract is a clouding of the lens of the eye due to light scattering of precipitated lens proteins or aberrant cellular debris. The major proteins in the lens are crystallins, and they are extensively deamidated during aging and cataracts. Deamidation has been detected at the domain and monomer interfaces of several crystallins during aging. The purpose of this study was to determine the effects of two potential deamidation sites at the predicted interface of the betaA3-crystallin dimer on its structure and stability. The glutamine residues at the reported in vivo deamidation sites of Q180 in the C-terminal domain and at the homologous site Q85 in the N-terminal domain were substituted with glutamic acid residues by site-directed mutagenesis. Far-UV and near-UV circular dichroism spectroscopy indicated that there were subtle differences in the secondary structure and more notable differences in the tertiary structure of the mutant proteins compared to that of the wild type betaA3-crystallin. The Q85E/Q180E mutant also was more susceptible to enzymatic digestion, suggesting increased solvent accessibility. These structural changes in the deamidated mutants led to decreased stability during unfolding in urea and increased precipitation during heat denaturation. When simulating deamidation at both residues, there was a further decrease in stability and loss of cooperativity. However, multiangle-light scattering and quasi-elastic light scattering experiments showed that dimer formation was not disrupted, nor did higher-order oligomers form. These results suggest that introducing charges at the predicted domain interface in the betaA3 homodimer may contribute to the insolubilization of lens crystallins or favor other, more stable, crystallin subunit interactions.
Figures



Similar articles
-
Deamidation alters interactions of beta-crystallins in hetero-oligomers.Mol Vis. 2009;15:241-9. Epub 2009 Jan 28. Mol Vis. 2009. PMID: 19190732 Free PMC article.
-
Deamidation destabilizes and triggers aggregation of a lens protein, betaA3-crystallin.Protein Sci. 2008 Sep;17(9):1565-75. doi: 10.1110/ps.035410.108. Epub 2008 Jun 20. Protein Sci. 2008. PMID: 18567786 Free PMC article.
-
Decreasing the homodimer interaction: a common mechanism shared by the deltaG91 mutation and deamidation in betaA3-crystallin.Mol Vis. 2010 Mar 16;16:438-44. Mol Vis. 2010. PMID: 20300566 Free PMC article.
-
Lens β-crystallins: the role of deamidation and related modifications in aging and cataract.Prog Biophys Mol Biol. 2014 Jul;115(1):21-31. doi: 10.1016/j.pbiomolbio.2014.02.004. Epub 2014 Mar 6. Prog Biophys Mol Biol. 2014. PMID: 24613629 Free PMC article. Review.
-
Chemical Properties Determine Solubility and Stability in βγ-Crystallins of the Eye Lens.Chembiochem. 2021 Apr 16;22(8):1329-1346. doi: 10.1002/cbic.202000739. Epub 2021 Feb 10. Chembiochem. 2021. PMID: 33569867 Free PMC article. Review.
Cited by
-
Structure-based prediction of asparagine and aspartate degradation sites in antibody variable regions.PLoS One. 2014 Jun 24;9(6):e100736. doi: 10.1371/journal.pone.0100736. eCollection 2014. PLoS One. 2014. PMID: 24959685 Free PMC article.
-
Multiple Aggregation Pathways in Human γS-Crystallin and Its Aggregation-Prone G18V Variant.Invest Ophthalmol Vis Sci. 2017 Apr 1;58(4):2397-2405. doi: 10.1167/iovs.16-20621. Invest Ophthalmol Vis Sci. 2017. PMID: 28444328 Free PMC article.
-
Cholesterol and Q147E Deamidation Modulates αA-Crystallin Membrane Binding Elucidating Protective Role of Lens Membrane Composition Changes With Aging.Invest Ophthalmol Vis Sci. 2025 May 1;66(5):8. doi: 10.1167/iovs.66.5.8. Invest Ophthalmol Vis Sci. 2025. PMID: 40323268 Free PMC article.
-
Aggregation of deamidated human betaB2-crystallin and incomplete rescue by alpha-crystallin chaperone.Exp Eye Res. 2010 Jun;90(6):688-98. doi: 10.1016/j.exer.2010.02.007. Epub 2010 Feb 23. Exp Eye Res. 2010. PMID: 20188088 Free PMC article.
-
Structural analysis of the mutant protein D26G of human γS-crystallin, associated with Coppock cataract.Mol Vis. 2013 Jun 5;19:1231-7. Print 2013. Mol Vis. 2013. PMID: 23761725 Free PMC article.
References
-
- Delaye M, Tardieu A. Short-range order of crystallin proteins accounts for eye lens transparency. Nature. 1983;302:415–417. - PubMed
-
- Zhang Z, Smith DL, Smith JB. Human beta-crystallins modified by backbone cleavage, deamidation and oxidation are prone to associate. Exp Eye Res. 2003;77:259–272. - PubMed
-
- Lapko VN, Smith DL, Smith JB. S-methylated cysteines in human lens gamma S-crystallins. Biochemistry. 2002;41:14645–14651. - PubMed
-
- Hanson SR, Hasan A, Smith DL, Smith JB. The major in vivo modifications of the human water-insoluble lens crystallins are disulfide bonds, deamidation, methionine oxidation and backbone cleavage. Exp Eye Res. 2000;71:195–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
