

The mutT defect does not elevate chromosomal fragmentation in Escherichia coli because of the surprisingly low levels of MutM/MutY-recognized DNA modifications
- PMID: 17616589
- PMCID: PMC2045204
- DOI: 10.1128/JB.00776-07
The mutT defect does not elevate chromosomal fragmentation in Escherichia coli because of the surprisingly low levels of MutM/MutY-recognized DNA modifications
Abstract
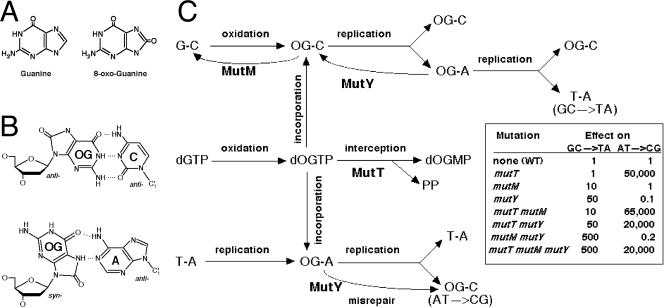
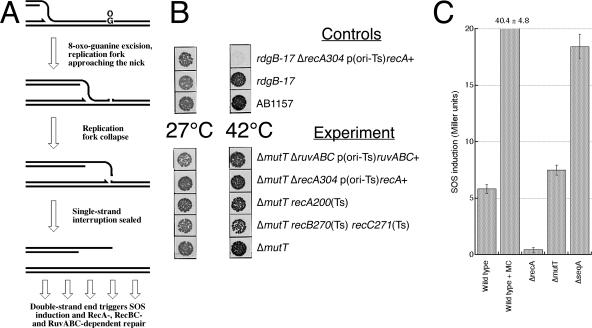
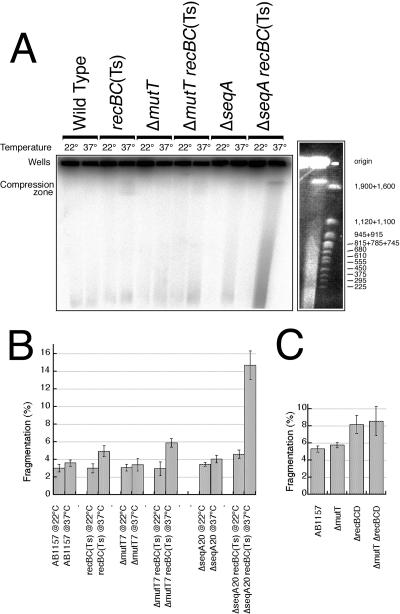
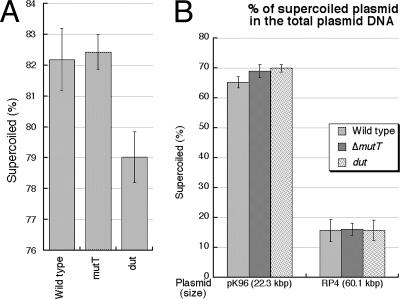
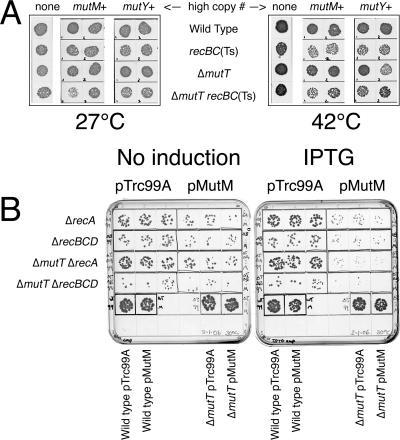
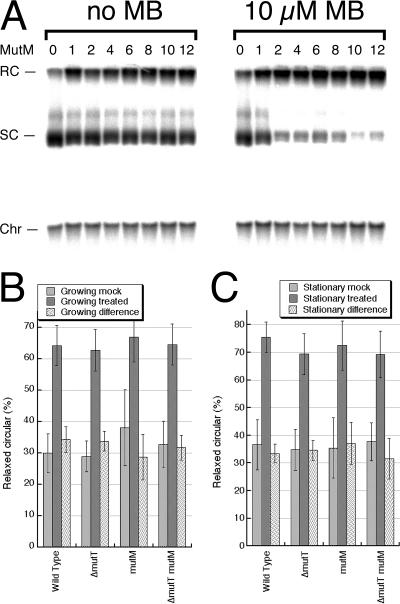
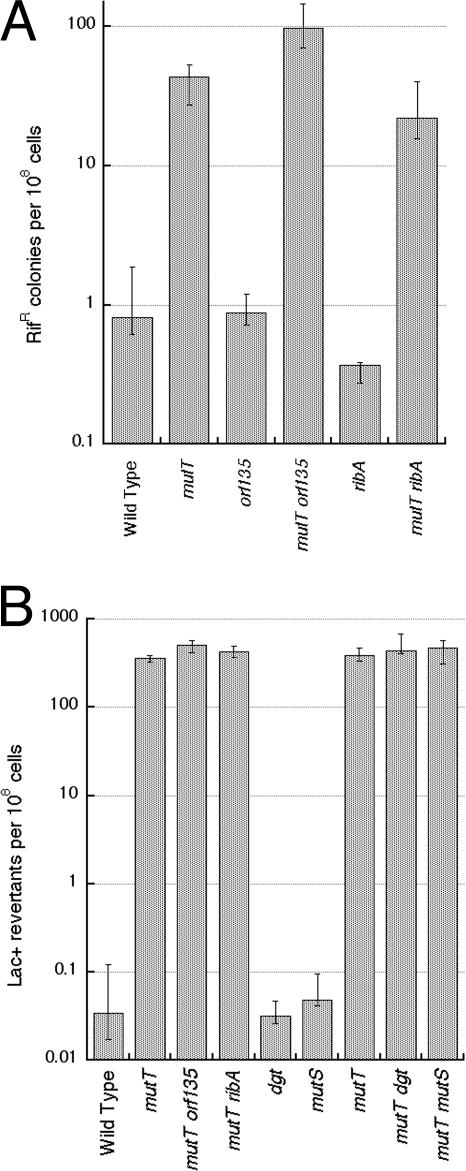
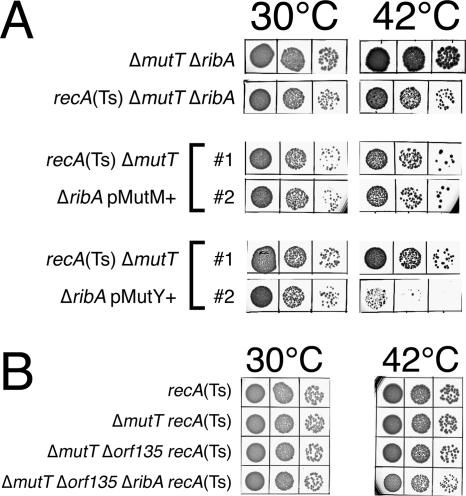
Nucleotide pool sanitizing enzymes Dut (dUTPase), RdgB (dITPase), and MutT (8-oxo-dGTPase) of Escherichia coli hydrolyze noncanonical DNA precursors to prevent incorporation of base analogs into DNA. Previous studies reported dramatic AT-->CG mutagenesis in mutT mutants, suggesting a considerable density of 8-oxo-G in DNA that should cause frequent excision and chromosomal fragmentation, irreparable in the absence of RecBCD-catalyzed repair and similar to the lethality of dut recBC and rdgB recBC double mutants. In contrast, we found mutT recBC double mutants viable with no signs of chromosomal fragmentation. Overproduction of the MutM and MutY DNA glycosylases, both acting on DNA containing 8-oxo-G, still yields no lethality in mutT recBC double mutants. Plasmid DNA, extracted from mutT mutM double mutant cells and treated with MutM in vitro, shows no increased relaxation, indicating no additional 8-oxo-G modifications. Our DeltamutT allele elevates the AT-->CG transversion rate 27,000-fold, consistent with published reports. However, the rate of AT-->CG transversions in our mutT(+) progenitor strain is some two orders of magnitude lower than in previous studies, which lowers the absolute rate of mutagenesis in DeltamutT derivatives, translating into less than four 8-oxo-G modifications per genome equivalent, which is too low to cause the expected effects. Introduction of various additional mutations in the DeltamutT strain or treatment with oxidative agents failed to increase the mutagenesis even twofold. We conclude that, in contrast to the previous studies, there is not enough 8-oxo-G in the DNA of mutT mutants to cause elevated excision repair that would trigger chromosomal fragmentation.
Figures
Similar articles
-
Chromosomal fragmentation in dUTPase-deficient mutants of Escherichia coli and its recombinational repair.Mol Microbiol. 2004 Mar;51(5):1279-95. doi: 10.1111/j.1365-2958.2003.03924.x. Mol Microbiol. 2004. PMID: 14982624
-
Functional cooperation of MutT, MutM and MutY proteins in preventing mutations caused by spontaneous oxidation of guanine nucleotide in Escherichia coli.Mutat Res. 1995 May;336(3):257-67. doi: 10.1016/0921-8777(94)00062-b. Mutat Res. 1995. PMID: 7739614
-
Analysis of spontaneous base substitutions generated in mutator strains of Bacillus subtilis.FEMS Microbiol Lett. 2004 May 1;234(1):37-42. doi: 10.1016/j.femsle.2004.03.004. FEMS Microbiol Lett. 2004. PMID: 15109717
-
Modulation of oxidative mutagenesis and carcinogenesis by polymorphic forms of human DNA repair enzymes.Mutat Res. 2005 Dec 11;591(1-2):60-73. doi: 10.1016/j.mrfmmm.2005.03.033. Epub 2005 Aug 2. Mutat Res. 2005. PMID: 16081110 Review.
-
MutT-related error avoidance mechanism for DNA synthesis.Genes Cells. 1996 Feb;1(2):139-45. doi: 10.1046/j.1365-2443.1996.d01-232.x. Genes Cells. 1996. PMID: 9140059 Review.
Cited by
-
Antibiotic killing through oxidized nucleotides.Proc Natl Acad Sci U S A. 2018 Feb 27;115(9):1967-1969. doi: 10.1073/pnas.1800255115. Epub 2018 Feb 14. Proc Natl Acad Sci U S A. 2018. PMID: 29444858 Free PMC article. No abstract available.
-
Cyanide, peroxide and nitric oxide formation in solutions of hydroxyurea causes cellular toxicity and may contribute to its therapeutic potency.J Mol Biol. 2009 Jul 31;390(5):845-62. doi: 10.1016/j.jmb.2009.05.038. Epub 2009 May 23. J Mol Biol. 2009. PMID: 19467244 Free PMC article.
-
Reduced lipopolysaccharide phosphorylation in Escherichia coli lowers the elevated ori/ter ratio in seqA mutants.Mol Microbiol. 2009 Jun;72(5):1273-92. doi: 10.1111/j.1365-2958.2009.06725.x. Epub 2009 Apr 30. Mol Microbiol. 2009. PMID: 19432803 Free PMC article.
-
Elevated Levels of DNA Strand Breaks Induced by a Base Analog in the Human Cell Line with the P32T ITPA Variant.J Nucleic Acids. 2010 Sep 26;2010:872180. doi: 10.4061/2010/872180. J Nucleic Acids. 2010. PMID: 20936128 Free PMC article.
-
Mutation rate dynamics in a bacterial population reflect tension between adaptation and genetic load.Proc Natl Acad Sci U S A. 2013 Jan 2;110(1):222-7. doi: 10.1073/pnas.1219574110. Epub 2012 Dec 17. Proc Natl Acad Sci U S A. 2013. PMID: 23248287 Free PMC article.
References
-
- Amado, L., and A. Kuzminov. 2006. The replication intermediates in Escherichia coli are not the product of DNA processing or uracil excision. J. Biol. Chem. 281:22635-22646. - PubMed
-
- Amann, E., B. Ochs, and K. J. Abel. 1988. Tightly regulated tac promoter vectors useful for the expression of unfused and fused proteins in Escherichia coli. Gene 69:301-315. - PubMed
-
- Bachmann, B. J. 1987. Derivations and genotypes of some mutant derivatives of Escherichia coli K-12, p. 1190-1219. In F. C. Neidhardt, L. Ingraham, K. B. Low, B. Magasanik, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella typhimurium: cellular and molecular biology. American Society for Microbiology, Washington, DC.
-
- Bertani, L. E., A. Haggmark, and P. Reichard. 1963. Enzymatic synthesis of deoxyribonucleotides. II. Formation and interconversion of deoxyuridine phosphates. J. Biol. Chem. 238:3407-3413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
