

Comment
doi: 10.1128/JB.00958-07.
Epub 2007 Jul 6.
Hypothetical functions of toxin-antitoxin systems
Affiliations
- PMID: 17616596
- PMCID: PMC1951896
- DOI: 10.1128/JB.00958-07
Item in Clipboard
Comment
Hypothetical functions of toxin-antitoxin systems
J Bacteriol.
2007 Sep.
No abstract available
Figures
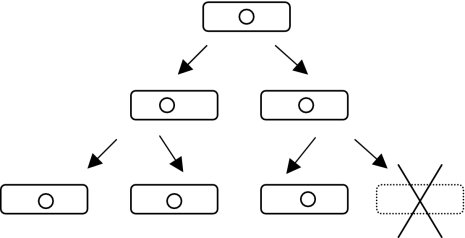
Toxin-antitoxin systems are addictive. An addictive genetic element (small circle) can increase effective plasmid stability approximately 10- to 1,000-fold (35) by the arrest or elimination of element-free daughter cells (lower right). Potentially, the loss of the addictive genetic element may occur by (mis-) segregration, exclusion, mutation, or recombination. If the environment is sufficiently structured and has limited resources, then the elimination of element-free cells will leave more food for element-containing cells. Since plasmid-borne toxin-antitoxin systems can be found on chromosomes (66) and since chromosomal toxin-antitoxin systems can stabilize plasmids (12, 26), there is at present little evidence of any obligatory location-specific specializations. Restriction modification systems (48), hok-sok-type postsegregational killing systems (25), and eukaryotic meiotic drive systems (6) possess similar addictive properties but are mechanistically distinct.
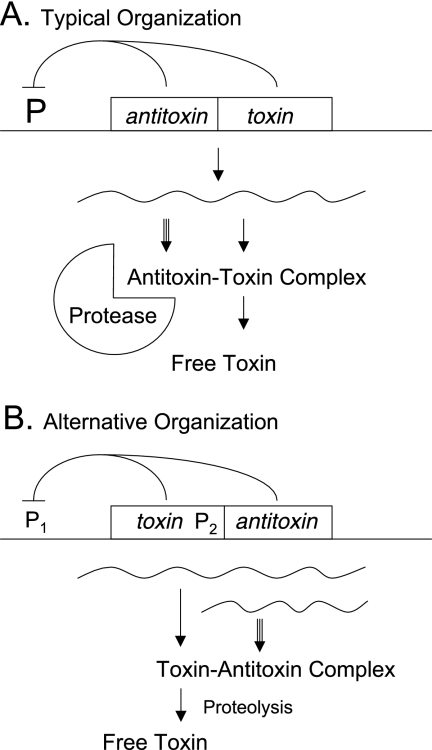
Organization of toxin-antitoxin systems. (A) Typically, the antitoxin gene precedes the toxin gene. (B) In some families, the usual order is reversed and a second promoter is observed. In both cases, antitoxin and toxin genes are usually small and often have overlapping stop and start codons, indicating that translation may be coupled. Transcription of the two genes is generally autoregulated by the protein products. The antitoxin is synthesized at a greater rate than the toxin but is also degraded at greater rate, due to the action of a specific host-encoded ATP-dependent protease, such as Lon, ClpXP, or ClpAP. At steady state, there is sufficient antitoxin to bind and neutralize the toxin. If the genes are lost or if their synthesis is sufficiently disrupted, the continuing proteolysis of the unstable antitoxin may liberate the stable toxin and produce biological effects. In at least a few atypical instances, a third protein product is implicated in the functioning of the system or the toxin is unusually large. Numerous reviews are available (19, 23, 29, 31, 35, 56, 57, 68, 70).

Phylogenetic network of chimeric toxin-antitoxin systems. The known toxins belong to a diverse and polyphyletic set of at least four major families, and the antitoxins are equally heterogeneous (4, 51). The N-terminal moiety of the antitoxin typically contains a DNA-binding domain belonging to any one of four of more repressor families, while the C-terminal moiety contains a flexible antitoxin domain that is typically too short and too diverse to be of any significant bioinformatic utility. The major toxin-antitoxin families can be loosely connected in an evolutionary sense only by their propensities to swap operator-repressor and antitoxin-toxin modules, thereby generating a network of chimeric systems (4, 22, 27, 30, 44, 63).
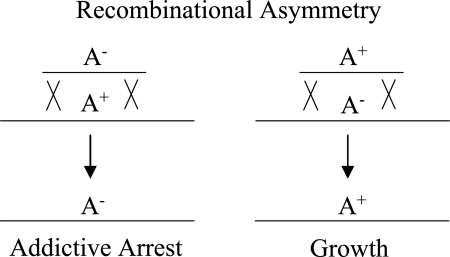
Recombinational asymmetry. The replacement of an addictive allele (A+) with a nonaddictive allele (A−) leads to addictive arrest, but the reciprocal process gives a viable recombinant. Thus, toxin-antitoxin systems and other addictive genetic elements can introduce an asymmetry into an otherwise symmetric gene transfer process, such as transduction, conjugation, or transformation. If the environment is structured and has limited resources, then the elimination of A− cells may leave more food for A+ cells and thus generate a slight benefit for the addictive allele as well as a slight detriment to the nonaddictive allele.
Comment on
-
What is the benefit to Escherichia coli of having multiple toxin-antitoxin systems in its genome?J Bacteriol. 2007 Sep;189(17):6101-8. doi: 10.1128/JB.00527-07. Epub 2007 May 18. J Bacteriol. 2007. PMID: 17513477 Free PMC article.
References
-
- Aguirre-Ramirez, M., J. Ramirez-Santos, L. Van Melderen, and M. C. Gomez-Eichelmann. 2006. Expression of the F plasmid ccd toxin-antitoxin system in Escherichia coli cells under nutritional stress. Can. J. Microbiol. 52:24-30. - PubMed
-
- Arcus, V. L., K. Backbro, A. Roos, E. L. Daniel, and E. N. Baker. 2004. Distant structural homology leads to the functional characterization of an archaeal PIN domain as an exonuclease. J. Biol. Chem. 279:16471-16478. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
