

LTR retrotransposons in rice (Oryza sativa, L.): recent burst amplifications followed by rapid DNA loss
- PMID: 17617907
- PMCID: PMC1940013
- DOI: 10.1186/1471-2164-8-218
LTR retrotransposons in rice (Oryza sativa, L.): recent burst amplifications followed by rapid DNA loss
Abstract
Background: LTR retrotransposons are one of the main causes for plant genome size and structure evolution, along with polyploidy. The characterization of their amplification and subsequent elimination of the genomes is therefore a major goal in plant evolutionary genomics. To address the extent and timing of these forces, we performed a detailed analysis of 41 LTR retrotransposon families in rice.
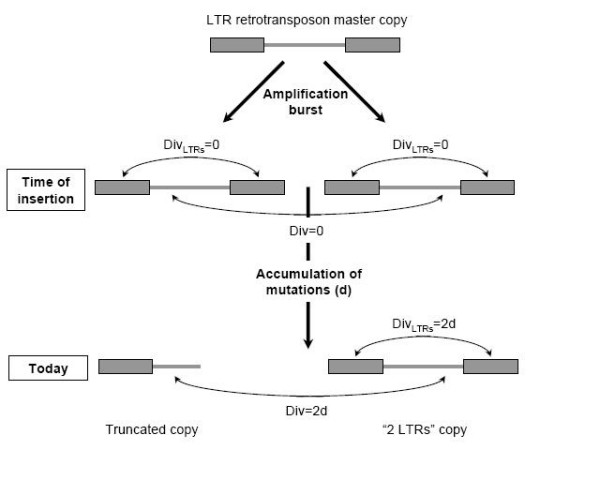
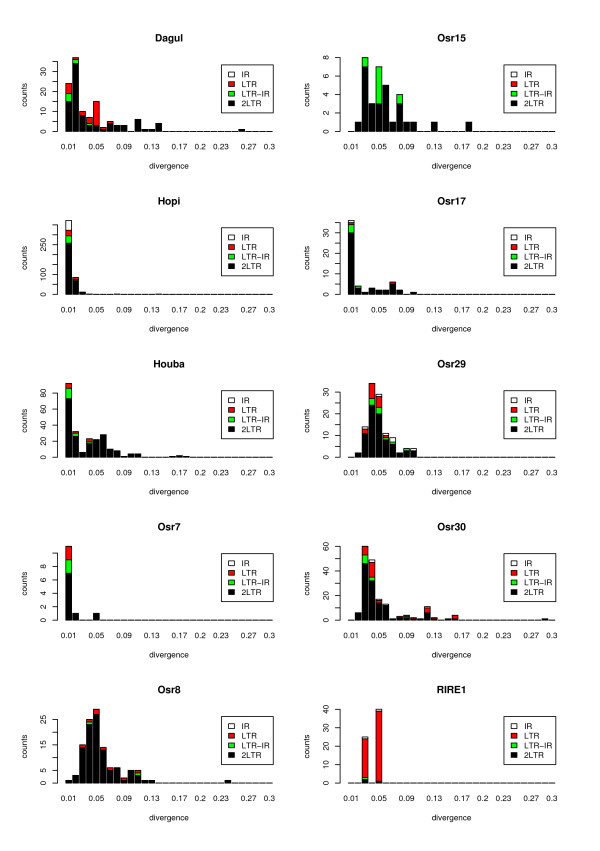
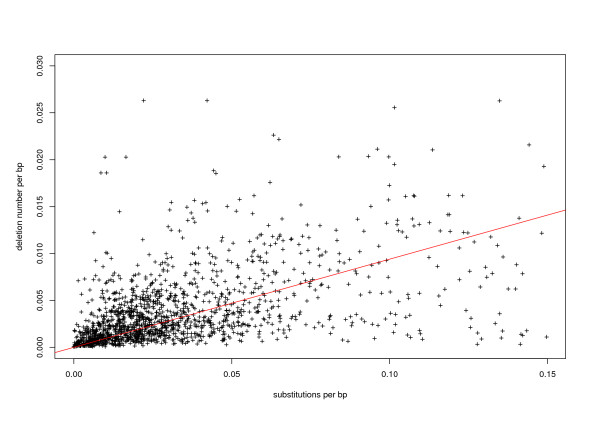
Results: Using a new method to estimate the insertion date of both truncated and complete copies, we estimated these two forces more accurately than previous studies based on other methods. We show that LTR retrotransposons have undergone bursts of amplification within the past 5 My. These bursts vary both in date and copy number among families, revealing that each family has a particular amplification history. The number of solo LTR varies among families and seems to correlate with LTR size, suggesting that solo LTR formation is a family-dependent process. The deletion rate estimate leads to the prediction that the half-life of LTR retrotransposon sequences evolving neutrally is about 19 My in rice, suggesting that other processes than the formation of small deletions are prevalent in rice DNA removal.
Conclusion: Our work provides insights into the dynamics of LTR retrotransposons in the rice genome. We show that transposable element families have distinct amplification patterns, and that the turn-over of LTR retrotransposons sequences is rapid in the rice genome.
Figures
References
-
- Kim JM, Vanguri S, Boeke JD, Gabriel A, Voytas DF. Transposable elements and genome organization: a comprehensive survey of retrotransposons revealed by the complete Saccharomyces cerevisiae genome sequence. Genome Res. 1998;8:464–78. - PubMed
-
- Kaminker JS, Bergman CM, Kronmiller B, Carlson J, Svirskas R, Patel S, Frise E, Wheeler DA, Lewis SE, Rubin GM, Ashburner M, Celniker SE. The transposable elements of the Drosophila melanogaster euchromatin: a genomics perspective. Genome Biol. 2002;3:RESEARCH0084. doi: 10.1186/gb-2002-3-12-research0084. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
