

Viral dsRNA inhibitors prevent self-association and autophosphorylation of PKR
- PMID: 17619024
- PMCID: PMC3710116
- DOI: 10.1016/j.jmb.2007.06.028
Viral dsRNA inhibitors prevent self-association and autophosphorylation of PKR
Abstract
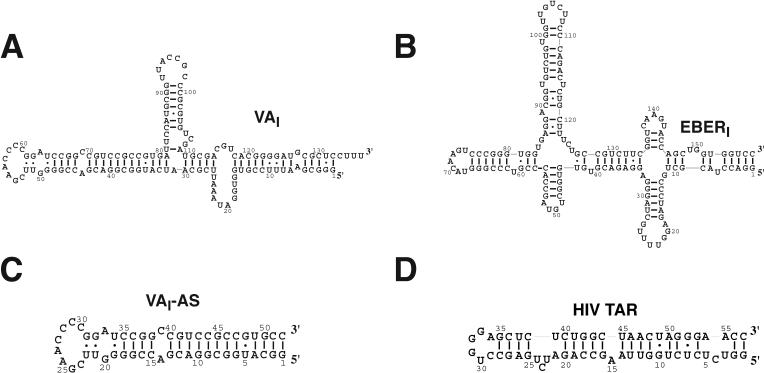
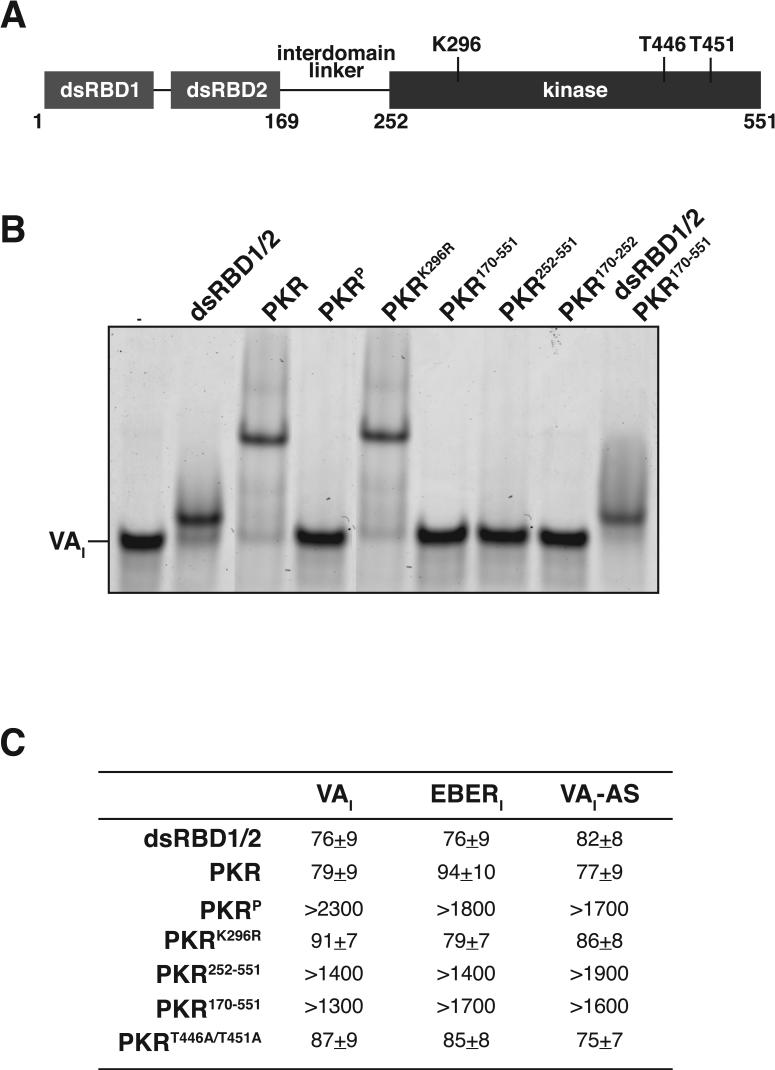
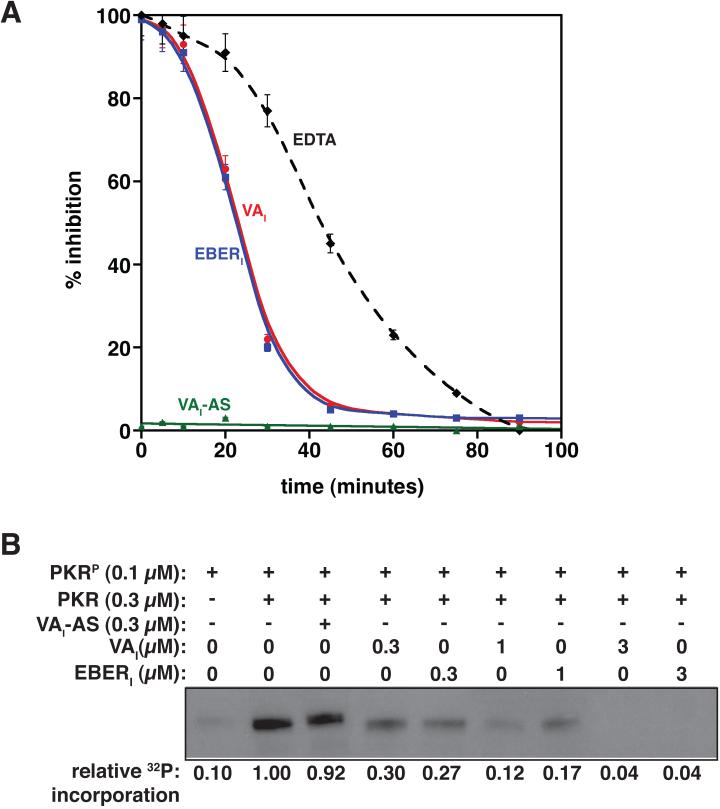
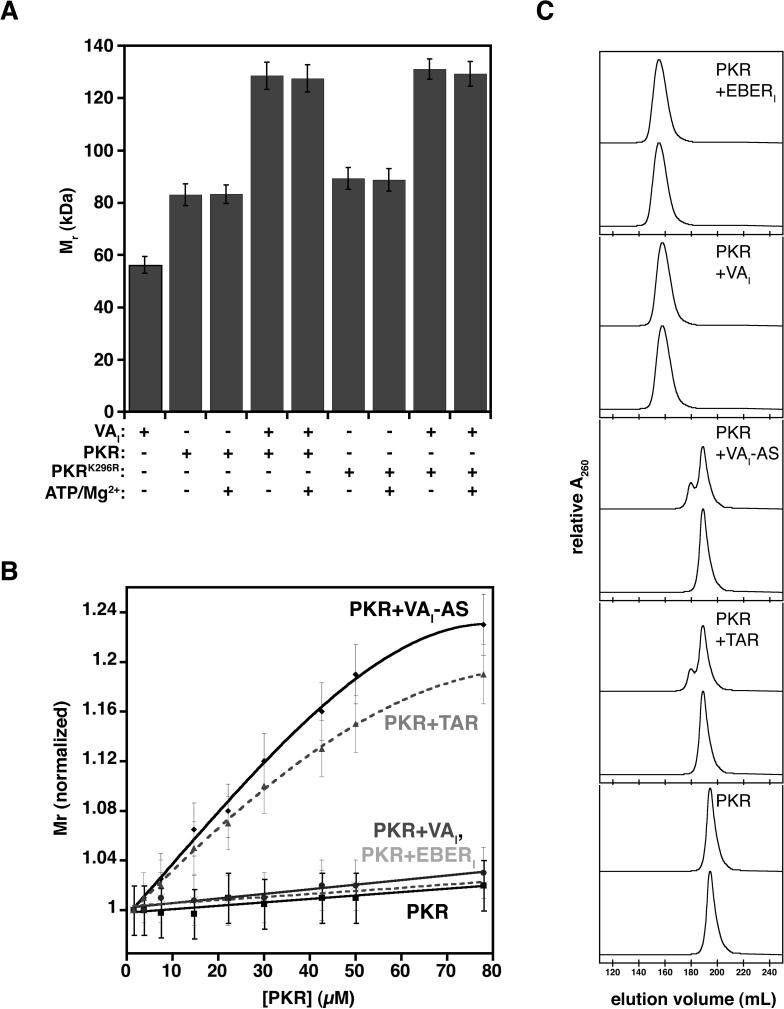
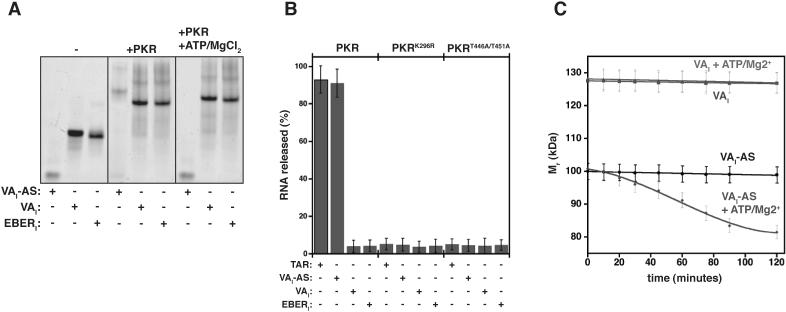
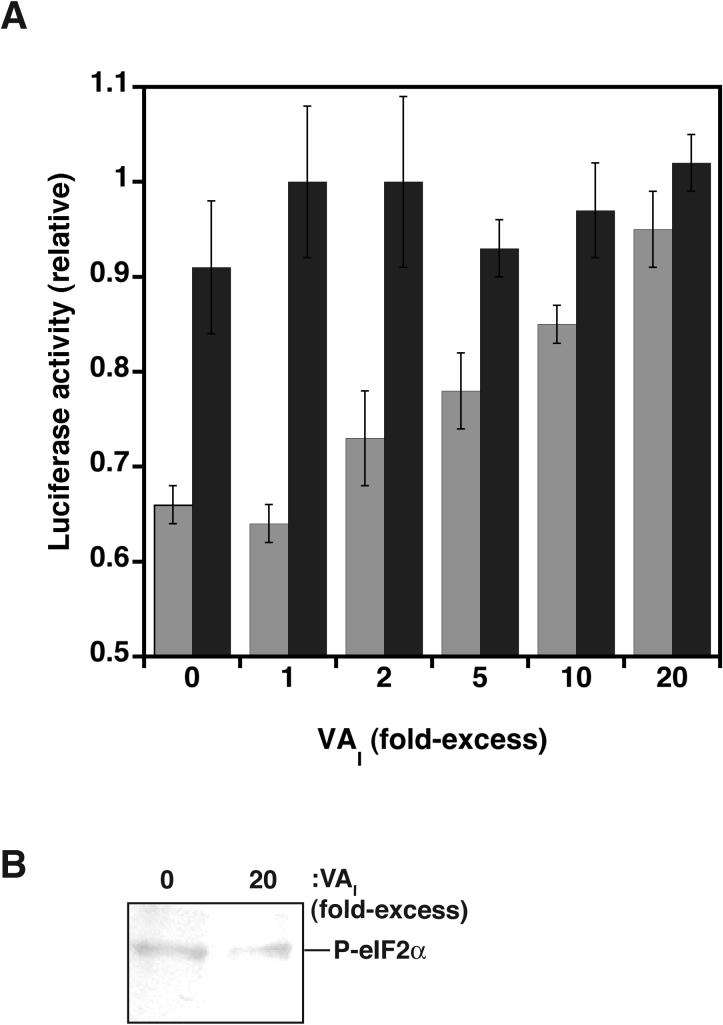
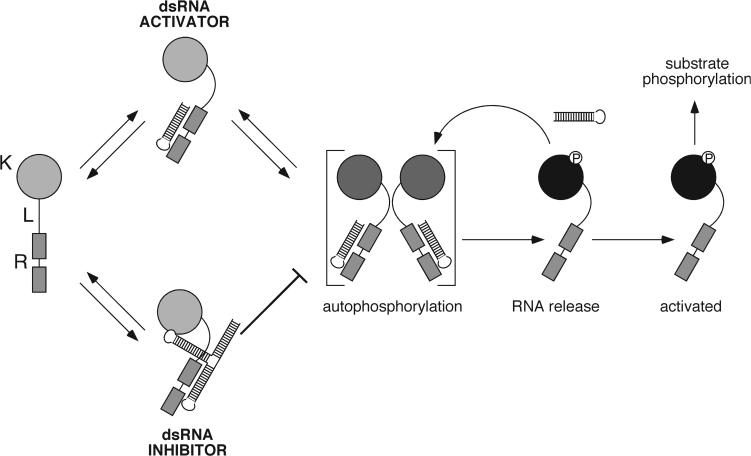
Host response to viral RNA genomes and replication products represents an effective strategy to combat viral invasion. PKR is a Ser/Thr protein kinase that binds to double-stranded (ds)RNA, autophosphorylates its kinase domain, and subsequently phosphorylates eukaryotic initiation factor 2alpha (eIF2alpha). This results in attenuation of protein translation, preventing synthesis of necessary viral proteins. In certain DNA viruses, PKR function can be evaded by transcription of highly structured virus-encoded dsRNA inhibitors that bind to and inactivate PKR. We probe here the mechanism of PKR inhibition by two viral inhibitor RNAs, EBER(I) (from Epstein-Barr) and VA(I) (from human adenovirus). Native gel shift mobility assays and isothermal titration calorimetry experiments confirmed that the RNA-binding domains of PKR are sufficient and necessary for the interaction with dsRNA inhibitors. Both EBER(I) and VA(I) are effective inhibitors of PKR activation by preventing trans-autophosphorylation between two PKR molecules. The RNA inhibitors prevent self-association of PKR molecules, providing a mechanistic basis for kinase inhibition. A variety of approaches indicated that dsRNA inhibitors remain associated with PKR under activating conditions, as opposed to activator dsRNA molecules that dissociate due to reduced affinity for the phosphorylated form of PKR. Finally, we show using a HeLa cell extract system that inhibitors of PKR result in translational recovery by the protein synthesis machinery. These data indicate that inhibitory dsRNAs bind preferentially to the latent, dephosphorylated form of PKR and prevent dimerization that is required for trans-autophosphorylation.
Figures
Similar articles
-
Uncoupling of RNA binding and PKR kinase activation by viral inhibitor RNAs.J Mol Biol. 2006 May 19;358(5):1270-85. doi: 10.1016/j.jmb.2006.03.003. Epub 2006 Mar 20. J Mol Biol. 2006. PMID: 16580685
-
Binding of double-stranded RNA to protein kinase PKR is required for dimerization and promotes critical autophosphorylation events in the activation loop.J Biol Chem. 2001 Jul 6;276(27):24946-58. doi: 10.1074/jbc.M102108200. Epub 2001 May 3. J Biol Chem. 2001. PMID: 11337501
-
RNA dimerization promotes PKR dimerization and activation.J Mol Biol. 2009 Jul 10;390(2):319-38. doi: 10.1016/j.jmb.2009.05.005. Epub 2009 May 13. J Mol Biol. 2009. PMID: 19445956 Free PMC article.
-
Regulation of innate immunity through RNA structure and the protein kinase PKR.Curr Opin Struct Biol. 2011 Feb;21(1):119-27. doi: 10.1016/j.sbi.2010.11.003. Epub 2010 Dec 8. Curr Opin Struct Biol. 2011. PMID: 21145228 Free PMC article. Review.
-
Activation of PKR: an open and shut case?Trends Biochem Sci. 2007 Feb;32(2):57-62. doi: 10.1016/j.tibs.2006.12.003. Epub 2006 Dec 29. Trends Biochem Sci. 2007. PMID: 17196820 Free PMC article. Review.
Cited by
-
The Structure-To-Function Relationships of Gammaherpesvirus-Encoded Long Non-Coding RNAs and Their Contributions to Viral Pathogenesis.Noncoding RNA. 2018 Sep 26;4(4):24. doi: 10.3390/ncrna4040024. Noncoding RNA. 2018. PMID: 30261651 Free PMC article. Review.
-
Mechanism of herpesvirus protein kinase UL13 in immune escape and viral replication.Front Immunol. 2022 Nov 30;13:1088690. doi: 10.3389/fimmu.2022.1088690. eCollection 2022. Front Immunol. 2022. PMID: 36531988 Free PMC article. Review.
-
Impact of the structural integrity of the three-way junction of adenovirus VAI RNA on PKR inhibition.PLoS One. 2017 Oct 20;12(10):e0186849. doi: 10.1371/journal.pone.0186849. eCollection 2017. PLoS One. 2017. PMID: 29053745 Free PMC article.
-
Specificity of the double-stranded RNA-binding domain from the RNA-activated protein kinase PKR for double-stranded RNA: insights from thermodynamics and small-angle X-ray scattering.Biochemistry. 2012 Nov 20;51(46):9312-22. doi: 10.1021/bi300935p. Epub 2012 Nov 9. Biochemistry. 2012. PMID: 23140277 Free PMC article.
-
Shaping the host cell environment with viral noncoding RNAs.Semin Cell Dev Biol. 2023 Sep 15;146:20-30. doi: 10.1016/j.semcdb.2022.12.008. Epub 2022 Dec 28. Semin Cell Dev Biol. 2023. PMID: 36581481 Free PMC article. Review.
References
-
- Malmgaard L. Induction and regulation of IFNs during viral infections. J Interferon Cytokine Res. 2004;24:439–54. - PubMed
-
- Peel AL. PKR activation in neurodegenerative disease. J Neuropathol Exp Neurol. 2004;63:97–105. - PubMed
-
- Langland JO, Cameron JM, Heck MC, Jancovich JK, Jacobs BL. Inhibition of PKR by RNA and DNA viruses. Virus Res. 2006;119:100–10. - PubMed
-
- Gale M,, Jr., Katze MG. Molecular mechanisms of interferon resistance mediated by viral-directed inhibition of PKR, the interferon-induced protein kinase. Pharmacol Ther. 1998;78:29–46. - PubMed
-
- Francois C, Duverlie G, Rebouillat D, Khorsi H, Castelain S, Blum HE, Gatignol A, Wychowski C, Moradpour D, Meurs EF. Expression of hepatitis C virus proteins interferes with the antiviral action of interferon independently of PKR-mediated control of protein synthesis. J Virol. 2000;74:5587–96. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
