

Axon viability and mitochondrial function are dependent on local protein synthesis in sympathetic neurons
- PMID: 17619140
- PMCID: PMC11517218
- DOI: 10.1007/s10571-007-9148-y
Axon viability and mitochondrial function are dependent on local protein synthesis in sympathetic neurons
Abstract
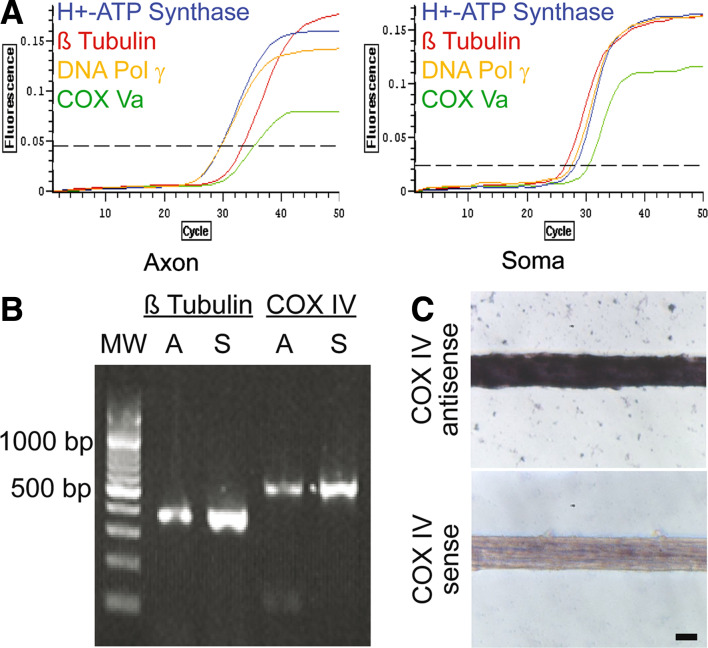
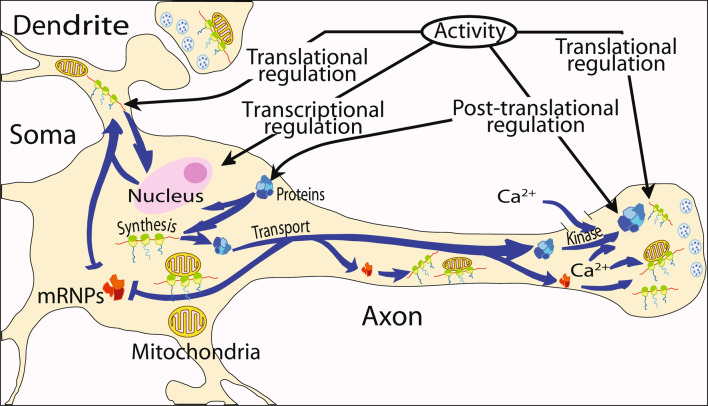
(1) Axons contain numerous mRNAs and a local protein synthetic system that can be regulated independently of the cell body. (2) In this study, cultured primary sympathetic neurons were employed, to assess the effect of local protein synthesis blockade on axon viability and mitochondrial function. (3) Inhibition of local protein synthesis reduced newly synthesized axonal proteins by 65% and resulted in axon retraction after 6 h. Acute inhibition of local protein synthesis also resulted in a significant decrease in the membrane potential of axonal mitochondria. Likewise, blockade of local protein transport into the mitochondria by transfection of the axons with Hsp90 C-terminal domain decreased the mitochondrial membrane potential by 65%. Moreover, inhibition of the local protein synthetic system also reduced the ability of mitochondria to restore axonal levels of ATP after KCl-induced depolarization. (4) Taken together, these results indicate that the local protein synthetic system plays an important role in mitochondrial function and the maintenance of the axon.
Figures



Similar articles
-
Intra-axonal synthesis of eukaryotic translation initiation factors regulates local protein synthesis and axon growth in rat sympathetic neurons.J Neurosci. 2013 Apr 24;33(17):7165-74. doi: 10.1523/JNEUROSCI.2040-12.2013. J Neurosci. 2013. PMID: 23616526 Free PMC article.
-
Axonal protein synthesis and the regulation of local mitochondrial function.Results Probl Cell Differ. 2009;48:225-42. doi: 10.1007/400_2009_1. Results Probl Cell Differ. 2009. PMID: 19343315 Free PMC article. Review.
-
AMP-activated protein kinase mediates activity-dependent axon branching by recruiting mitochondria to axon.Dev Neurobiol. 2014 Jun;74(6):557-73. doi: 10.1002/dneu.22149. Epub 2013 Nov 29. Dev Neurobiol. 2014. PMID: 24218086
-
Local translation of ATP synthase subunit 9 mRNA alters ATP levels and the production of ROS in the axon.Mol Cell Neurosci. 2012 Mar;49(3):263-70. doi: 10.1016/j.mcn.2011.12.006. Epub 2011 Dec 21. Mol Cell Neurosci. 2012. PMID: 22209705
-
Crosstalk of Local Translation and Mitochondria: Powering Plasticity in Axons and Dendrites.Neuron. 2019 Jan 16;101(2):204-206. doi: 10.1016/j.neuron.2018.12.027. Neuron. 2019. PMID: 30653934 Review.
Cited by
-
Mitochondrial dynamics regulate growth cone motility, guidance, and neurite growth rate in perinatal retinal ganglion cells in vitro.Invest Ophthalmol Vis Sci. 2012 Oct 30;53(11):7402-11. doi: 10.1167/iovs.12-10298. Invest Ophthalmol Vis Sci. 2012. PMID: 23049086 Free PMC article.
-
Axonal mRNA in uninjured and regenerating cortical mammalian axons.J Neurosci. 2009 Apr 15;29(15):4697-707. doi: 10.1523/JNEUROSCI.6130-08.2009. J Neurosci. 2009. PMID: 19369540 Free PMC article.
-
Making scent of the presence and local translation of odorant receptor mRNAs in olfactory axons.Dev Neurobiol. 2014 Mar;74(3):259-68. doi: 10.1002/dneu.22122. Epub 2013 Oct 7. Dev Neurobiol. 2014. PMID: 23959692 Free PMC article. Review.
-
Looking for answers far away from the soma-the (un)known axonal functions of TDP-43, and their contribution to early NMJ disruption in ALS.Mol Neurodegener. 2023 May 31;18(1):35. doi: 10.1186/s13024-023-00623-6. Mol Neurodegener. 2023. PMID: 37259156 Free PMC article. Review.
-
Mitochondrial fusion/fission, transport and autophagy in Parkinson's disease: when mitochondria get nasty.Parkinsons Dis. 2011 Feb 20;2011:767230. doi: 10.4061/2011/767230. Parkinsons Dis. 2011. PMID: 21403911 Free PMC article.
References
-
- Anderson S, Bankier AT, Barrell BG, de Bruijn MHL, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJH, Staden R, Young IG (1981) Sequence and organization of the human mitochondrial genome. Nature 290:457–465 - PubMed
-
- Attardi G, Schatz G (1988) Biogenesis of mitochondria. Annu Rev Cell Biol 4:289–331 - PubMed
-
- Bauer MF, Hofmann S (2006) Import of mitochondrial proteins. In: Schapira AHV (ed) Mitochondrial function and dysfunction. Academic Press, San Diego, CA, pp 57–90
-
- Beaumont V, Zhong N, Fletcher R, Froemke RC, Zucker RS (2001) Phosphorylation and local presynaptic protein synthesis in calcium- and calcineurin-dependent induction of crayfish long-term facilitation. Neuron 32:489–501 - PubMed
-
- Bleher R, Martin R (2001) Ribosomes in the squid giant axon. Neuroscience 107:527–534 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
