

Cdk8 is essential for preimplantation mouse development
- PMID: 17620419
- PMCID: PMC1952144
- DOI: 10.1128/MCB.01302-06
Cdk8 is essential for preimplantation mouse development
Abstract
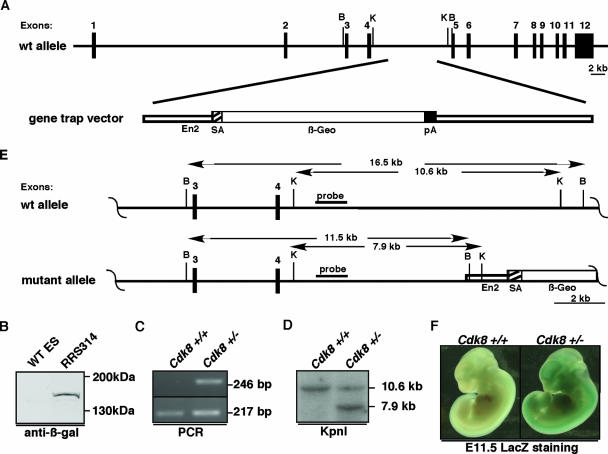
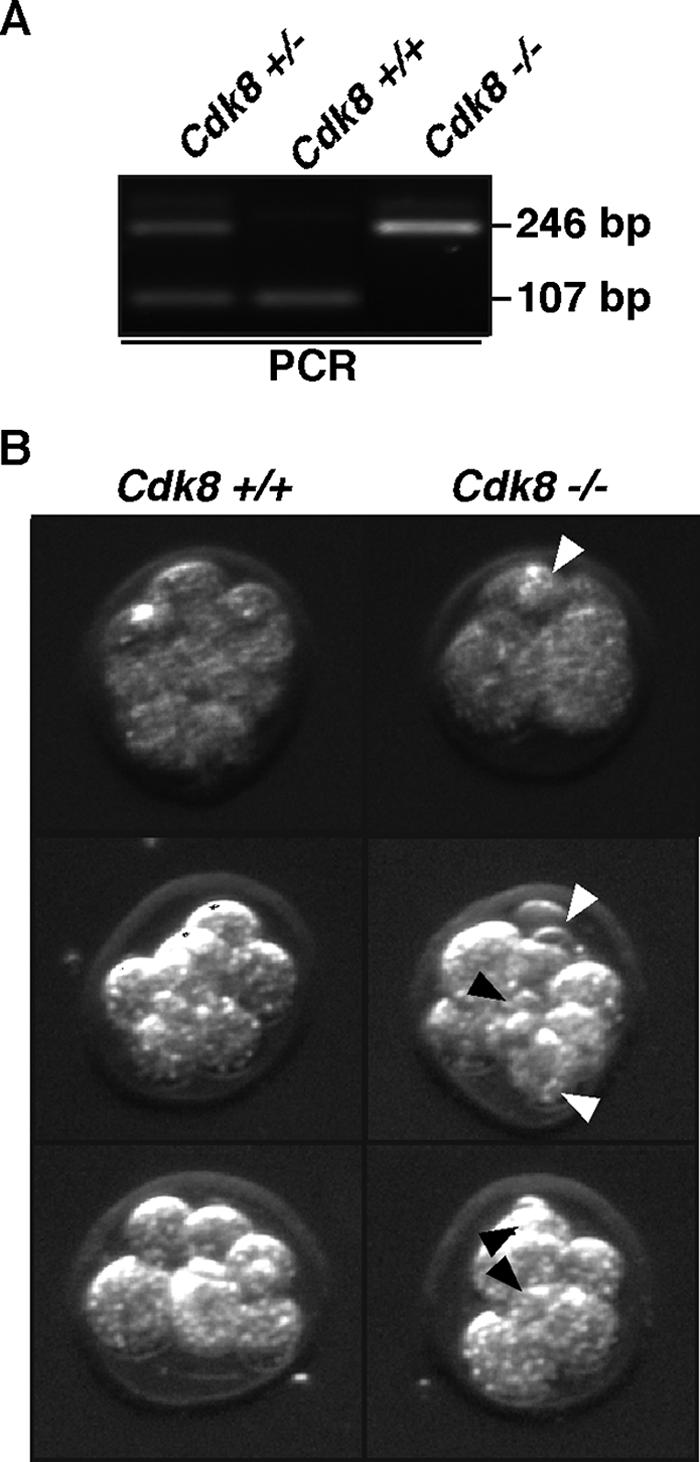
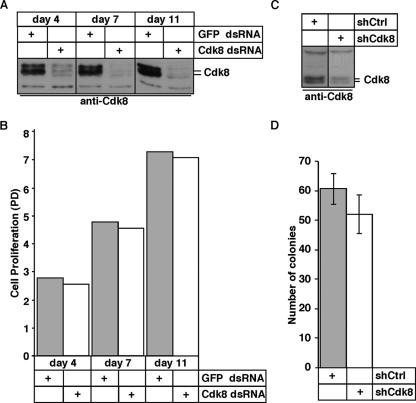
The Cdk8 kinase and associated proteins form a nonessential transcriptional repressor module of the Mediator in the budding yeast Saccharomyces cerevisiae. Genetic analyses of this module have demonstrated functions ranging from environmental responses in budding yeast to organogenesis and development in worms, flies, and zebrafish. Here we have investigated the function of mammalian Cdk8 using mice harboring a gene trap insertion at the Cdk8 locus inactivating this kinase. No phenotypes were noted in heterozygote Cdk8+/- mice, but intercrossing these did not produce homozygous Cdk8-/- offspring. Developmental analysis demonstrated a requirement for Cdk8 prior to implantation at embryonic days 2.5 to 3.0. Cdk8-/- preimplantation embryos had fragmented blastomeres and did not proceed to compaction. As Cdk8 deficiency in cultured metazoan cells did not affect cell viability, the results suggest that transcriptional repression of genes critical for early-cell-fate determination underlies the requirement of Cdk8 in embryogenesis.
Figures
References
-
- Bettencourt-Dias, M., R. Giet, R. Sinka, A. Mazumdar, W. G. Lock, F. Balloux, P. J. Zafiropoulos, S. Yamaguchi, S. Winter, R. W. Carthew, M. Cooper, D. Jones, L. Frenz, and D. M. Glover. 2004. Genome-wide survey of protein kinases required for cell cycle progression. Nature 432:980-987. - PubMed
-
- Bjorklund, M., M. Taipale, M. Varjosalo, J. Saharinen, J. Lahdenpera, and J. Taipale. 2006. Identification of pathways regulating cell size and cell-cycle progression by RNAi. Nature 439:1009-1013. - PubMed
-
- Bjorklund, S., and C. M. Gustafsson. 2005. The yeast Mediator complex and its regulation. Trends Biochem. Sci. 30:240-244. - PubMed
-
- Borggrefe, T., R. Davis, H. Erdjument-Bromage, P. Tempst, and R. D. Kornberg. 2002. A complex of the Srb8, -9, -10, and -11 transcriptional regulatory proteins from yeast. J. Biol. Chem. 277:44202-44207. - PubMed
-
- Brummelkamp, T. R., R. Bernards, and R. Agami. 2002. Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell 2:243-247. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials