

Transcription factor NF-kappaB is transported to the nucleus via cytoplasmic dynein/dynactin motor complex in hippocampal neurons
- PMID: 17622342
- PMCID: PMC1899224
- DOI: 10.1371/journal.pone.0000589
Transcription factor NF-kappaB is transported to the nucleus via cytoplasmic dynein/dynactin motor complex in hippocampal neurons
Abstract
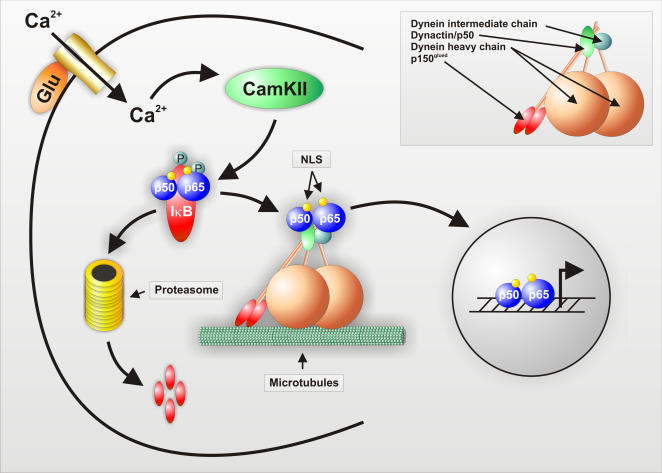
Background: Long-term changes in synaptic plasticity require gene transcription, indicating that signals generated at the synapse must be transported to the nucleus. Synaptic activation of hippocampal neurons is known to trigger retrograde transport of transcription factor NF-kappaB. Transcription factors of the NF-kappaB family are widely expressed in the nervous system and regulate expression of several genes involved in neuroplasticity, cell survival, learning and memory.
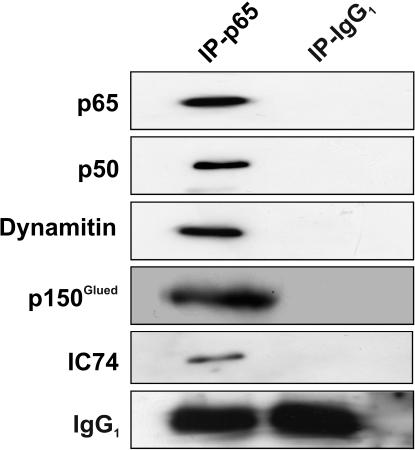
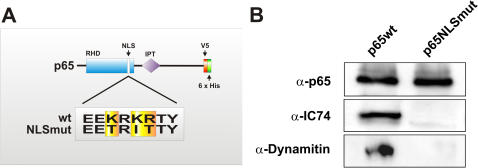
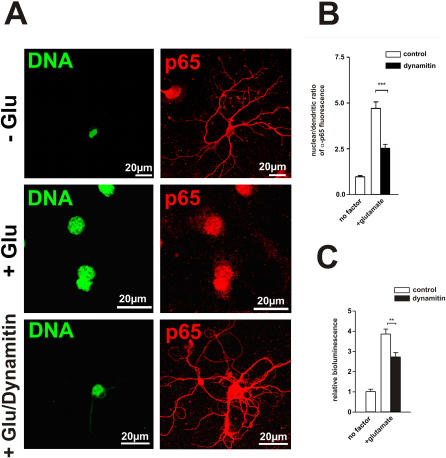
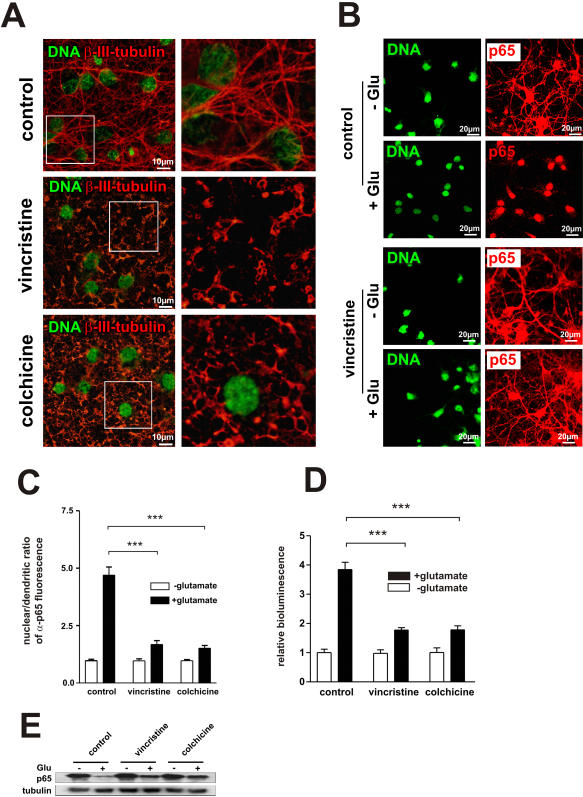
Principal findings: In this study, we examine the role of the dynein/dynactin motor complex in the cellular mechanism targeting and transporting activated NF-kappaB to the nucleus in response to synaptic stimulation. We demonstrate that overexpression of dynamitin, which is known to dissociate dynein from microtubules, and treatment with microtubule-disrupting drugs inhibits nuclear accumulation of NF-kappaB p65 and reduces NF-kappaB-dependent transcription activity. In this line, we show that p65 is associated with components of the dynein/dynactin complex in vivo and in vitro and that the nuclear localization sequence (NLS) within NF-kappaB p65 is essential for this binding.
Conclusion: This study shows the molecular mechanism for the retrograde transport of activated NF-kappaB from distant synaptic sites towards the nucleus.
Conflict of interest statement
Figures
References
-
- Bonizzi G, Karin M. The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004;25:280–288. - PubMed
-
- Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell 109 Suppl. 2002:S81–96. - PubMed
-
- Karin M, Lin A. NF-kappaB at the crossroads of life and death. Nat Immunol. 2002;3:221–227. - PubMed
-
- Li Q, Verma IM. NF-kappaB regulation in the immune system. Nat Rev Immunol. 2002;2:725–734. - PubMed
-
- Dingwall C, Laskey RA. Nuclear targeting sequences–a consensus? Trends Biochem Sci. 1991;16:478–481. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
