

Orang-like manual adaptations in the fossil hominoid Hispanopithecus laietanus: first steps towards great ape suspensory behaviours
- PMID: 17623642
- PMCID: PMC2274979
- DOI: 10.1098/rspb.2007.0750
Orang-like manual adaptations in the fossil hominoid Hispanopithecus laietanus: first steps towards great ape suspensory behaviours
Abstract
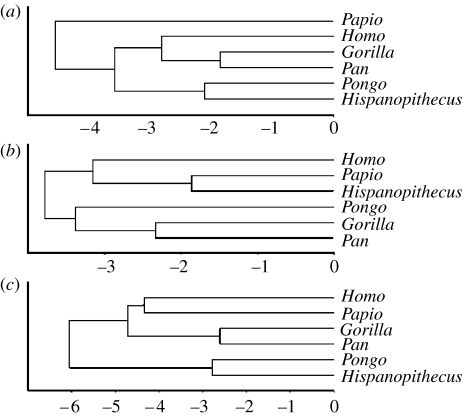
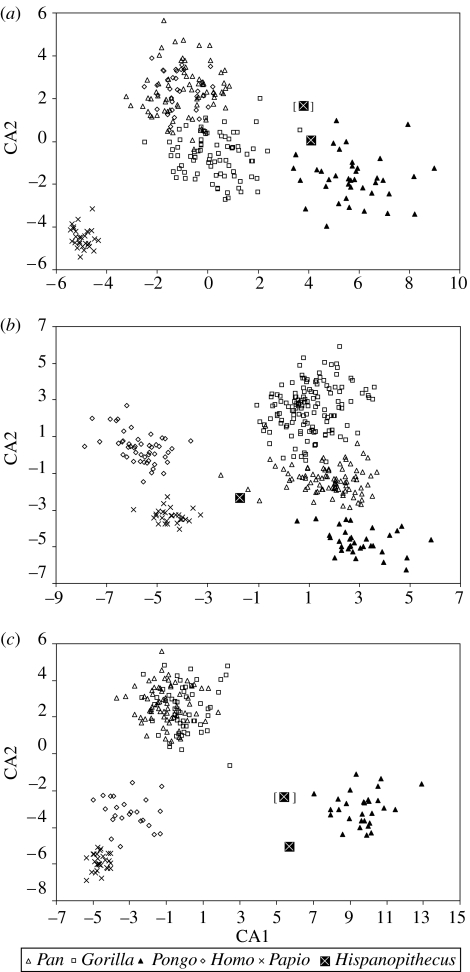
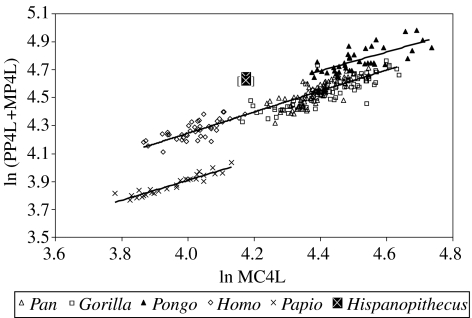
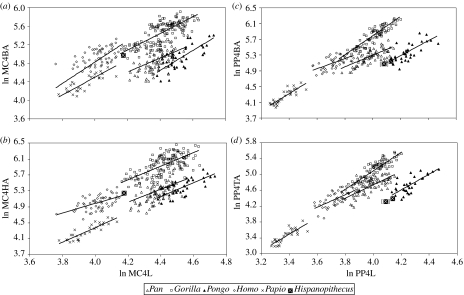
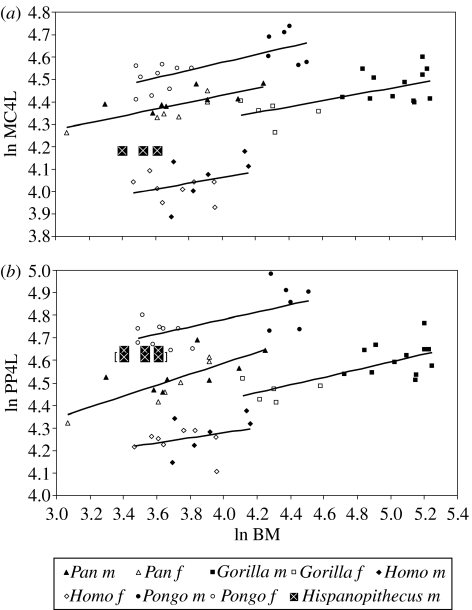
Morphological and biometrical analyses of the partial hand IPS18800 of the fossil great ape Hispanopithecus laietanus (=Dryopithecus laietanus), from the Late Miocene (about 9.5Ma) of Can Llobateres (Catalonia, Spain), reveal many similarities with extant orang-utans (Pongo). These similarities are interpreted as adaptations to below-branch suspensory behaviours, including arm-swinging and clambering/postural feeding on slender arboreal supports, due to an orang-like double-locking mechanism. This is confirmed by the long and highly curved phalanges of Hispanopithecus. The short and stout metacarpals with dorsally constricted heads, together with the dorsally extended articular facets on proximal phalanges, indicate the persistence of significant degrees of palmigrady. A powerful grasping capability is indicated by the great development of basal phalangeal tubercles, the marked insertions for the flexors on phalangeal shafts and the large pits for the collateral ligaments. The morphology of the Hispanopithecus long bones of the hand indicates a unique positional repertoire, combining orthogrady with suspensory behaviours and palmigrade quadrupedalism. The retention of powerful grasping and palmigrady suggests that the last common ancestor of hominids might have been more primitive than what can be inferred on the basis of extant taxa, suggesting that pronograde behaviours are compatible with an orthograde bodyplan suitable for climbing and suspension.
Figures
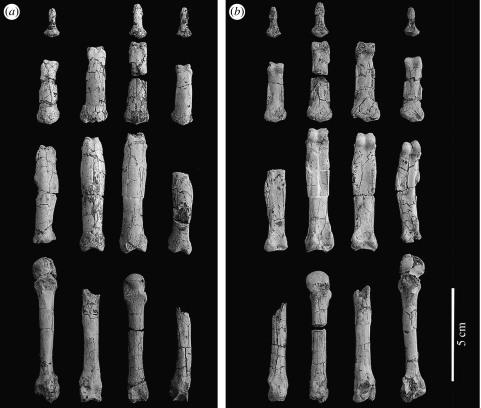
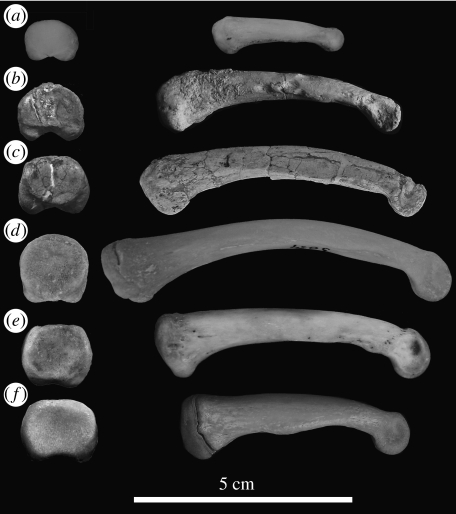
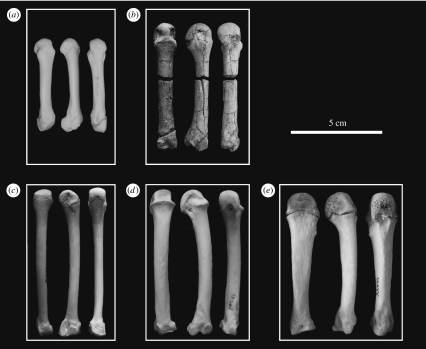
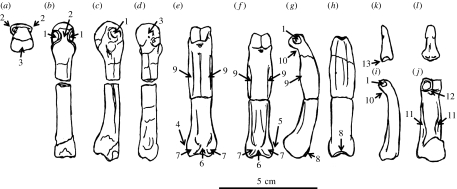
Comment in
-
Comment: an early ape shows its hand.Proc Biol Sci. 2007 Oct 7;274(1624):2373-4. doi: 10.1098/rspb.2007.0855. Proc Biol Sci. 2007. PMID: 17686726 Free PMC article. No abstract available.
References
-
- Alba D.M, Moyà-Solà S, Köhler M. Morphological affinities of the Australopithecus afarensis hand on the basis of manual proportions and relative thumb length. J. Hum. Evol. 2003;44:225–254. doi:10.1016/S0047-2484(02)00207-5 - DOI - PubMed
-
- Begun D.R. New catarrhine phalanges from Rudabánya (Northeastern Hungary) and the problem of parallelism and convergence in hominoid postcranial morphology. J. Hum. Evol. 1993;24:373–402. doi:10.1006/jhev.1993.1028 - DOI
-
- Begun D.R, Teaford M.F, Walker A.C. Comparative and functional anatomy of Proconsul phalanges from the Kaswanga Primate Site, Rusinga Island, Kenya. J. Hum. Evol. 1994;26:89–165. doi:10.1006/jhev.1994.1008 - DOI
-
- Begun D.R, Ward C.V, Rose M.D. Events in hominoid evolution. In: Begun D.R, Ward C.V, Rose M.D, editors. Function, phylogeny and fossils: miocene hominoid evolution and adaptation. Plenum Press; New York, NY: 1997. pp. 389–415.
-
- Gould S.J. Allometry and size in ontogeny and phylogeny. Biol. Rev. 1966;41:587–640. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
