

Tomato MAPKs LeMPK1, LeMPK2, and LeMPK3 function in the systemin-mediated defense response against herbivorous insects
- PMID: 17623784
- PMCID: PMC1924534
- DOI: 10.1073/pnas.0700344104
Tomato MAPKs LeMPK1, LeMPK2, and LeMPK3 function in the systemin-mediated defense response against herbivorous insects
Abstract
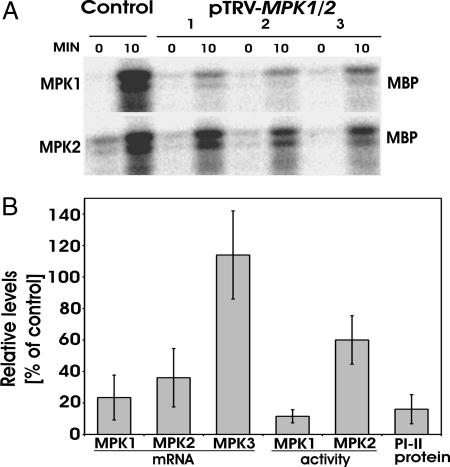
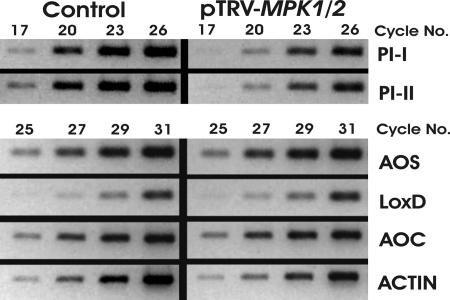
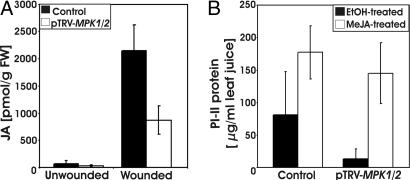
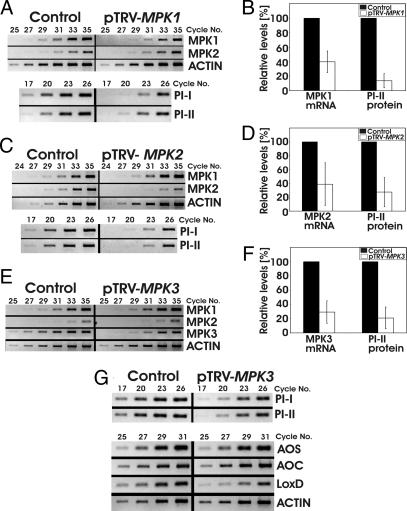
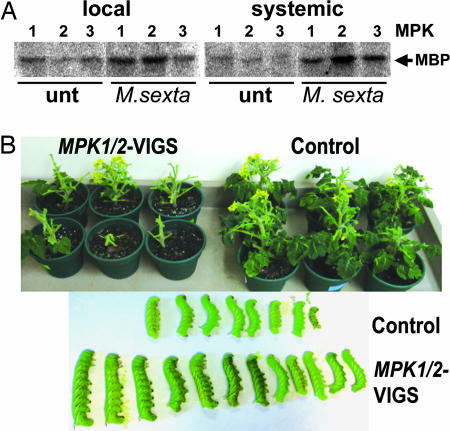
Systemin is a wound-signaling peptide that mediates defenses of tomato plants against herbivorous insects. Perception of systemin by the membrane-bound receptor SR160 results in activation of MAPKs, synthesis of jasmonic acid (JA), and expression of defense genes. To test the function of MAPKs in the response to systemin, we used virus-induced gene silencing (VIGS) in plants that overexpress the systemin precursor prosystemin (35S::prosys plants). These transgenic plants accumulate high levels of defense proteins and exhibit increased resistance to herbivorous insects. Cosilencing of the MAPKs MPK1 and MPK2 reduced MPK1/2 kinase activity, JA biosynthesis, and expression of JA-dependent defense genes. Application of methyl-JA restored the full defense response. These data show that MPK1 and MPK2 are essential components of the systemin signaling pathway and most likely function upstream of JA biosynthesis. MPK1 and MPK2 are 95% identical at the amino acid level. Specific VIGS of only MPK1 or MPK2 resulted in the same reduction of defense gene expression as cosilencing of MPK1 and MPK2, indicating that gene dosage effects may be important for MPK signaling. In addition, VIGS of the closely related MPK3 also reduced systemin-induced defense responses. The function of MPK1/2 and orthologs in pathogen-induced defenses is well established. Here we show that cosilencing of MPK1 and MPK2 compromised prosystemin-mediated resistance to Manduca sexta (Lepidoptera) herbivory, demonstrating that MPK1 and MPK2 are also required for successful defenses against herbivorous insects.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The tomato suppressor of prosystemin-mediated responses2 gene encodes a fatty acid desaturase required for the biosynthesis of jasmonic acid and the production of a systemic wound signal for defense gene expression.Plant Cell. 2003 Jul;15(7):1646-61. doi: 10.1105/tpc.012237. Plant Cell. 2003. PMID: 12837953 Free PMC article.
-
Systemin and jasmonic acid regulate constitutive and herbivore-induced systemic volatile emissions in tomato, Solanum lycopersicum.Phytochemistry. 2010 Dec;71(17-18):2024-37. doi: 10.1016/j.phytochem.2010.09.010. Epub 2010 Oct 21. Phytochemistry. 2010. PMID: 20970815
-
The tomato mutant spr1 is defective in systemin perception and the production of a systemic wound signal for defense gene expression.Plant J. 2003 Feb;33(3):567-76. doi: 10.1046/j.1365-313x.2003.01646.x. Plant J. 2003. PMID: 12581314
-
Systemin/Jasmonate-mediated systemic defense signaling in tomato.Mol Plant. 2011 Jul;4(4):607-15. doi: 10.1093/mp/ssr008. Epub 2011 Feb 28. Mol Plant. 2011. PMID: 21357647 Review.
-
The wound response in tomato--role of jasmonic acid.J Plant Physiol. 2006 Feb;163(3):297-306. doi: 10.1016/j.jplph.2005.10.014. Epub 2005 Dec 20. J Plant Physiol. 2006. PMID: 16368162 Review.
Cited by
-
Potato StMPK7 is a downstream component of StMKK1 and promotes resistance to the oomycete pathogen Phytophthora infestans.Mol Plant Pathol. 2021 Jun;22(6):644-657. doi: 10.1111/mpp.13050. Epub 2021 Mar 25. Mol Plant Pathol. 2021. PMID: 33764635 Free PMC article.
-
Opposite roles of MAPKKK17 and MAPKKK21 against Tetranychus urticae in Arabidopsis.Front Plant Sci. 2022 Dec 7;13:1038866. doi: 10.3389/fpls.2022.1038866. eCollection 2022. Front Plant Sci. 2022. PMID: 36570948 Free PMC article.
-
Tomato PEPR1 ORTHOLOG RECEPTOR-LIKE KINASE1 Regulates Responses to Systemin, Necrotrophic Fungi, and Insect Herbivory.Plant Cell. 2018 Sep;30(9):2214-2229. doi: 10.1105/tpc.17.00908. Epub 2018 Aug 21. Plant Cell. 2018. PMID: 30131419 Free PMC article.
-
A Disease Resistance Elicitor Laminarin Enhances Tea Defense against a Piercing Herbivore Empoasca (Matsumurasca) onukii Matsuda.Sci Rep. 2019 Jan 28;9(1):814. doi: 10.1038/s41598-018-37424-7. Sci Rep. 2019. PMID: 30692583 Free PMC article.
-
Survey of Sensitivity to Fatty Acid-Amino Acid Conjugates in the Solanaceae.J Chem Ecol. 2020 Mar;46(3):330-343. doi: 10.1007/s10886-020-01152-y. Epub 2020 Jan 27. J Chem Ecol. 2020. PMID: 31989490
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
