

Inhibition of HIV-1 replication by P-TEFb inhibitors DRB, seliciclib and flavopiridol correlates with release of free P-TEFb from the large, inactive form of the complex
- PMID: 17625008
- PMCID: PMC1948018
- DOI: 10.1186/1742-4690-4-47
Inhibition of HIV-1 replication by P-TEFb inhibitors DRB, seliciclib and flavopiridol correlates with release of free P-TEFb from the large, inactive form of the complex
Abstract
Background: The positive transcription elongation factor, P-TEFb, comprised of cyclin dependent kinase 9 (Cdk9) and cyclin T1, T2 or K regulates the productive elongation phase of RNA polymerase II (Pol II) dependent transcription of cellular and integrated viral genes. P-TEFb containing cyclin T1 is recruited to the HIV long terminal repeat (LTR) by binding to HIV Tat which in turn binds to the nascent HIV transcript. Within the cell, P-TEFb exists as a kinase-active, free form and a larger, kinase-inactive form that is believed to serve as a reservoir for the smaller form.
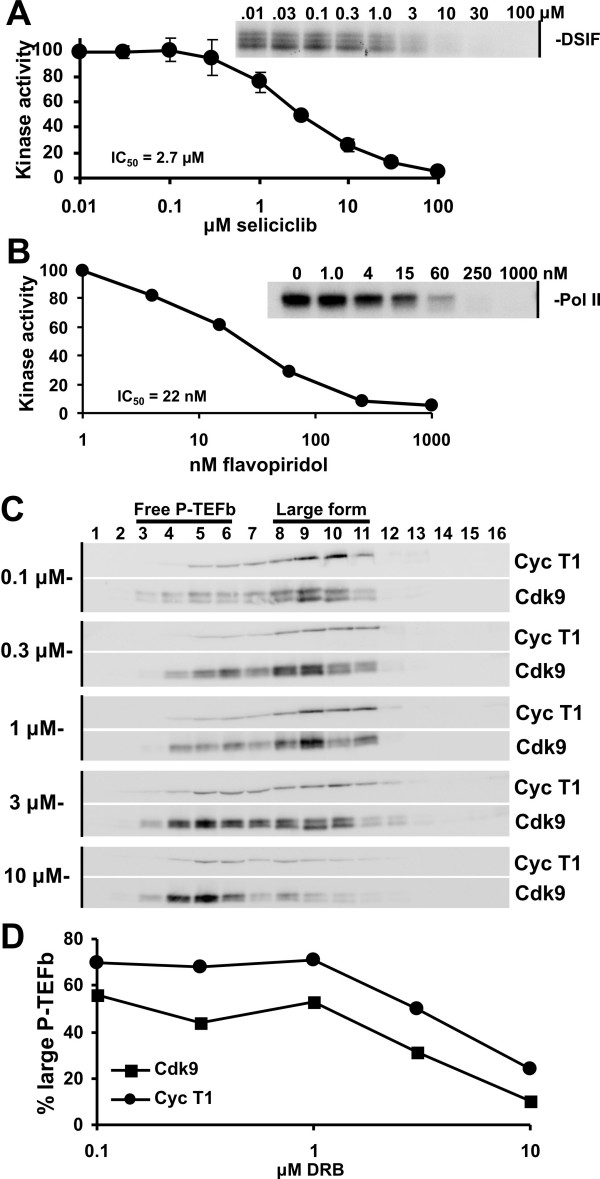
Results: We developed a method to rapidly quantitate the relative amounts of the two forms based on differential nuclear extraction. Using this technique, we found that titration of the P-TEFb inhibitors flavopiridol, DRB and seliciclib onto HeLa cells that support HIV replication led to a dose dependent loss of the large form of P-TEFb. Importantly, the reduction in the large form correlated with a reduction in HIV-1 replication such that when 50% of the large form was gone, HIV-1 replication was reduced by 50%. Some of the compounds were able to effectively block HIV replication without having a significant impact on cell viability. The most effective P-TEFb inhibitor flavopiridol was evaluated against HIV-1 in the physiologically relevant cell types, peripheral blood lymphocytes (PBLs) and monocyte derived macrophages (MDMs). Flavopiridol was found to have a smaller therapeutic index (LD50/IC50) in long term HIV-1 infectivity studies in primary cells due to greater cytotoxicity and reduced efficacy at blocking HIV-1 replication.
Conclusion: Initial short term studies with P-TEFb inhibitors demonstrated a dose dependent loss of the large form of P-TEFb within the cell and a concomitant reduction in HIV-1 infectivity without significant cytotoxicity. These findings suggested that inhibitors of P-TEFb may serve as effective anti-HIV-1 therapies. However, longer term HIV-1 replication studies indicated that these inhibitors were more cytotoxic and less efficacious against HIV-1 in the primary cell cultures.
Figures





References
-
- Fujinaga K, Irwin D, Huang Y, Taube R, Kurosu T, Peterlin BM. Dynamics of human immunodeficiency virus transcription: P-TEFb phosphorylates RD and dissociates negative effectors from the transactivation response element. Mol Cell Biol. 2004;24:787–795. doi: 10.1128/MCB.24.2.787-795.2004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
