

An intracellular P2X receptor required for osmoregulation in Dictyostelium discoideum
- PMID: 17625565
- PMCID: PMC3942652
- DOI: 10.1038/nature05926
An intracellular P2X receptor required for osmoregulation in Dictyostelium discoideum
Abstract
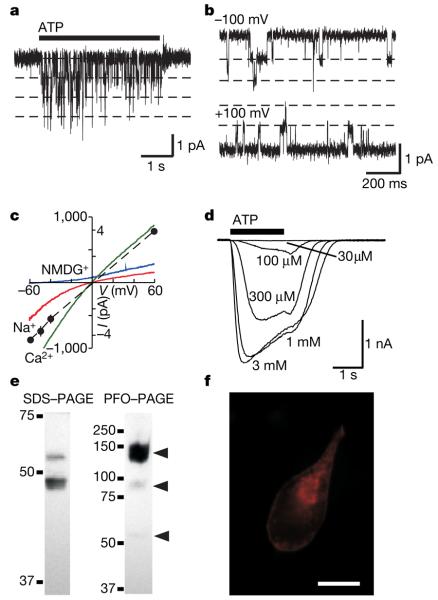
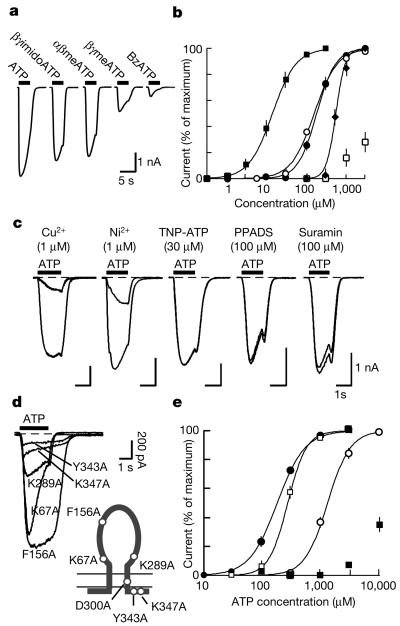
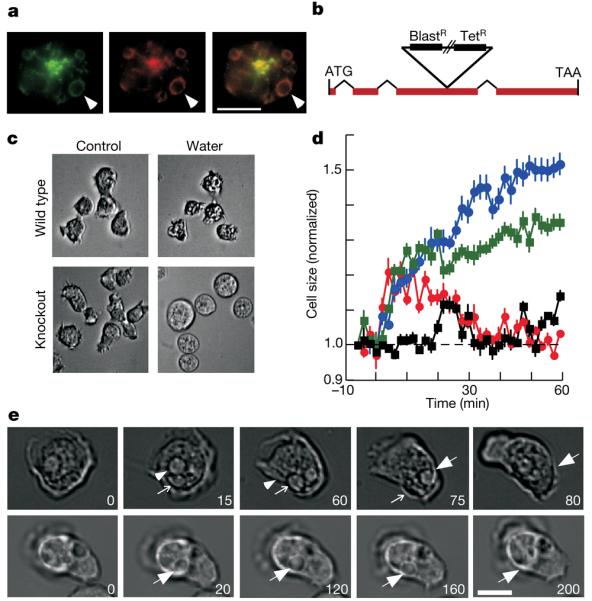
P2X receptors are membrane ion channels gated by extracellular ATP that are found widely in vertebrates, but not previously in microbes. Here we identify a weakly related gene in the genome of the social amoeba Dictyostelium discoideum, and show, with the use of heterologous expression in human embryonic kidney cells, that it encodes a membrane ion channel activated by ATP (30-100 muM). Site-directed mutagenesis revealed essential conservation of structure-function relations with P2X receptors of higher organisms. The receptor was insensitive to the usual P2X antagonists but was blocked by nanomolar concentrations of Cu2+ ions. In D. discoideum, the receptor was found on intracellular membranes, with prominent localization to an osmoregulatory organelle, the contractile vacuole. Targeted disruption of the gene in D. discoideum resulted in cells that were unable to regulate cell volume in hypotonic conditions. Cell swelling in these mutant cells was accompanied by a marked inhibition of contractile vacuole emptying. These findings demonstrate a new functional role for P2X receptors on intracellular organelles, in this case in osmoregulation.
Figures
Similar articles
-
Functional characterization of intracellular Dictyostelium discoideum P2X receptors.J Biol Chem. 2009 Dec 11;284(50):35227-39. doi: 10.1074/jbc.M109.045674. Epub 2009 Oct 15. J Biol Chem. 2009. PMID: 19833731 Free PMC article.
-
Intracellular P2X receptors as novel calcium release channels and modulators of osmoregulation in Dictyostelium: a comparison of two common laboratory strains.Channels (Austin). 2013 Jan 1;7(1):43-6. doi: 10.4161/chan.22737. Epub 2012 Dec 6. Channels (Austin). 2013. PMID: 23221467 Free PMC article.
-
Functional properties of five Dictyostelium discoideum P2X receptors.J Biol Chem. 2013 Jul 19;288(29):20992-21000. doi: 10.1074/jbc.M112.445346. Epub 2013 Jun 5. J Biol Chem. 2013. PMID: 23740252 Free PMC article.
-
Signaling at purinergic P2X receptors.Annu Rev Physiol. 2009;71:333-59. doi: 10.1146/annurev.physiol.70.113006.100630. Annu Rev Physiol. 2009. PMID: 18851707 Review.
-
Molecular physiology of P2X receptors.Physiol Rev. 2002 Oct;82(4):1013-67. doi: 10.1152/physrev.00015.2002. Physiol Rev. 2002. PMID: 12270951 Review.
Cited by
-
Ancient origin of four-domain voltage-gated Na+ channels predates the divergence of animals and fungi.J Membr Biol. 2012 Feb;245(2):117-23. doi: 10.1007/s00232-012-9415-9. Epub 2012 Jan 19. J Membr Biol. 2012. PMID: 22258316
-
Multiple roles of the extracellular vestibule amino acid residues in the function of the rat P2X4 receptor.PLoS One. 2013;8(3):e59411. doi: 10.1371/journal.pone.0059411. Epub 2013 Mar 21. PLoS One. 2013. PMID: 23555667 Free PMC article.
-
Epithelial wound healing in Clytia hemisphaerica provides insights into extracellular ATP signaling mechanisms and P2XR evolution.Sci Rep. 2023 Nov 1;13(1):18819. doi: 10.1038/s41598-023-45424-5. Sci Rep. 2023. PMID: 37914720 Free PMC article.
-
P2X4 forms functional ATP-activated cation channels on lysosomal membranes regulated by luminal pH.J Biol Chem. 2014 Jun 20;289(25):17658-67. doi: 10.1074/jbc.M114.552158. Epub 2014 May 9. J Biol Chem. 2014. PMID: 24817123 Free PMC article.
-
A two-pore channel protein required for regulating mTORC1 activity on starvation.BMC Biol. 2020 Jan 22;18(1):8. doi: 10.1186/s12915-019-0735-4. BMC Biol. 2020. PMID: 31969153 Free PMC article.
References
-
- North RA. Molecular physiology of P2X receptors. Physiol. Rev. 2002;82:1013–1067. - PubMed
-
- Khakh BS, North RA. P2X receptors as cell-surface ATP sensors in health and disease. Nature. 2006;442:527–532. - PubMed
-
- Gever JR, Cockayne DA, Dillon MP, Burnstock G, Ford AP. Pharmacology of P2X channels. Pflügers Arch. 2006;452:513–537. - PubMed
-
- Barrera NP, Ormond SJ, Henderson RM, Murrell-Lagnado RD, Edwardson JM. Atomic force microscopy imaging demonstrates that P2X2 receptors are trimers but that P2X6 receptor subunits do not oligomerize. J. Biol. Chem. 2005;280:10759–10765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
