

Zinc fingers 1 and 7 of yeast TFIIIA are essential for assembly of a functional transcription complex on the 5 S RNA gene
- PMID: 17626045
- PMCID: PMC1950542
- DOI: 10.1093/nar/gkm517
Zinc fingers 1 and 7 of yeast TFIIIA are essential for assembly of a functional transcription complex on the 5 S RNA gene
Abstract
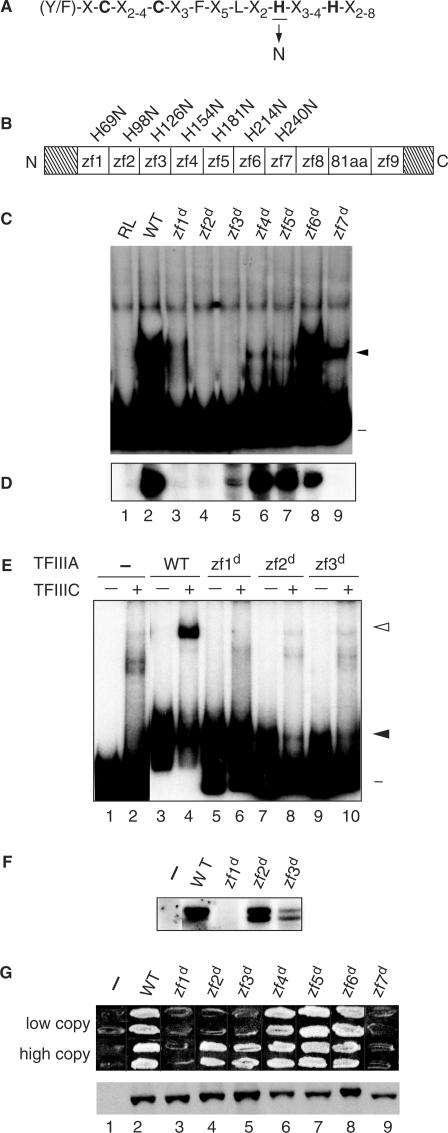
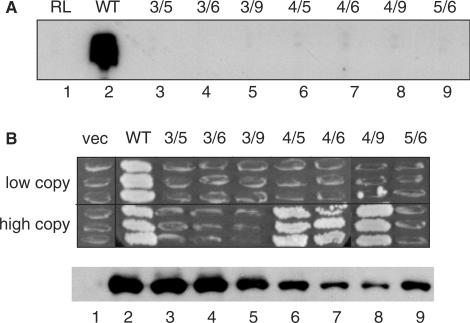
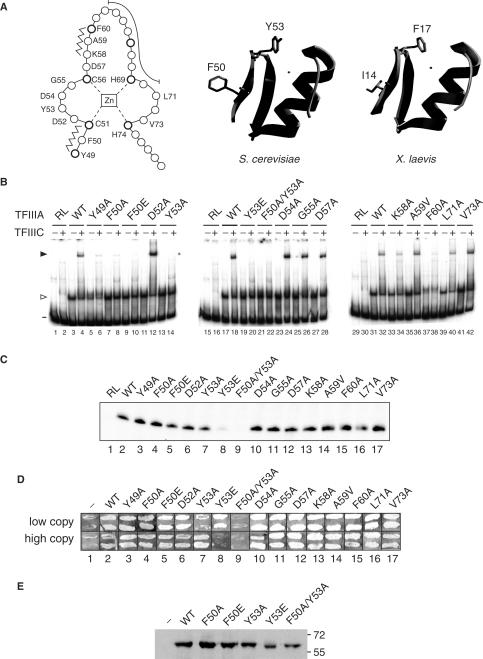
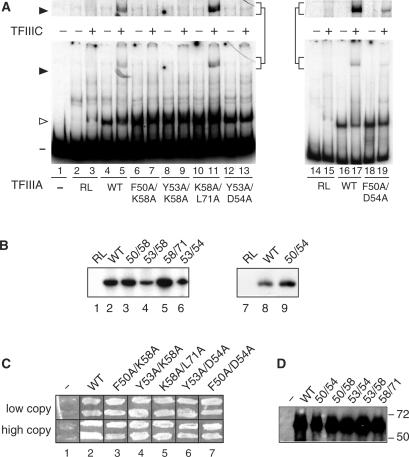
The binding of transcription factor (TF) IIIA to the internal control region of the 5 S RNA gene is the first step in the assembly of a DNA-TFIIIA-TFIIIC- TFIIIB transcription complex, which promotes accurate transcription by RNA polymerase III. With the use of mutations that are predicted to disrupt the folding of a zinc finger, we have examined the roles of zinc fingers 1 through 7 of yeast TFIIIA in the establishment of a functional transcription complex both in vitro and in vivo. Our data indicate that, in addition to their role in DNA binding, the first and seventh zinc fingers contribute other essential roles in the assembly of an active transcription complex. Alanine-scanning mutagenesis identified residues within zinc finger 1 that are not required for DNA binding but are required for incorporation of TFIIIC into the TFIIIA-DNA complex. Although disruption of zinc finger 2 or 3 had a deleterious effect on the activity of TFIIIA both in vitro and in vivo, we found that increasing the level of their in vivo expression allowed these mutant proteins to support cell viability. Disruption of zinc fingers 4, 5 or 6 had minimal effect on the DNA binding and TF activities of TFIIIA.
Figures
References
-
- Geiduschek EP, Kassavetis GA. RNA polymerase III transcription complexes. In: McKnight SL, Yamamoto KR, editors. Transcriptional Regulation. Vol. 1. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1992. pp. 247–280.
-
- White RJ. RNA Polymerase III Transcription. Austin: R. G. Landes Company; 1994.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
