

Regulation of the Arabidopsis root vascular initial population by LONESOME HIGHWAY
- PMID: 17626058
- PMCID: PMC3145339
- DOI: 10.1242/dev.006296
Regulation of the Arabidopsis root vascular initial population by LONESOME HIGHWAY
Abstract
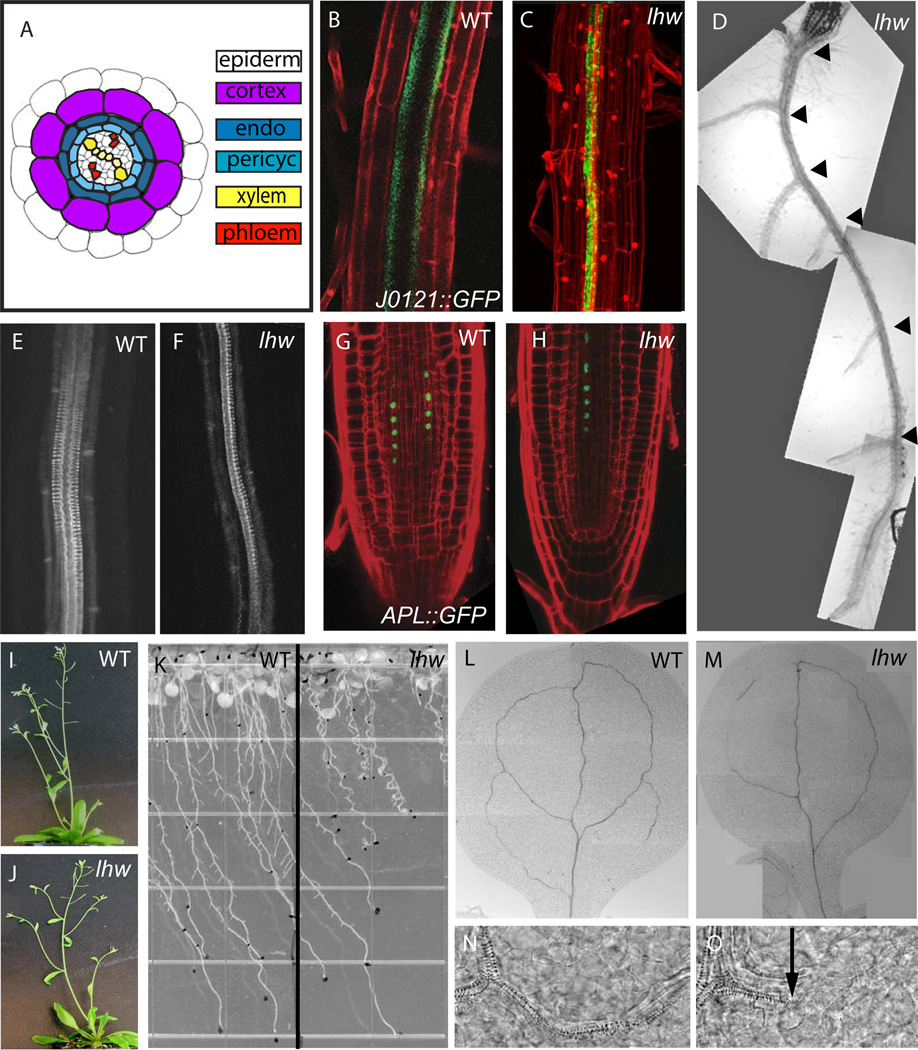
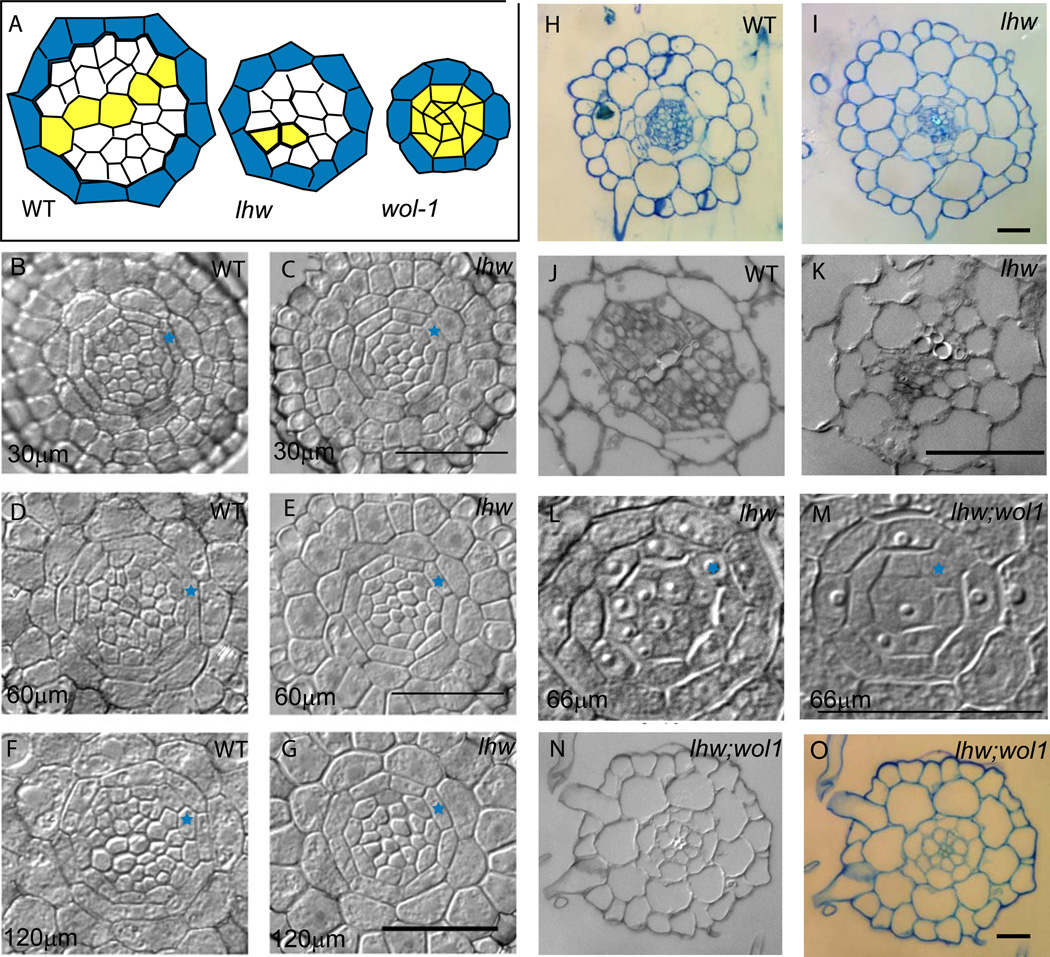
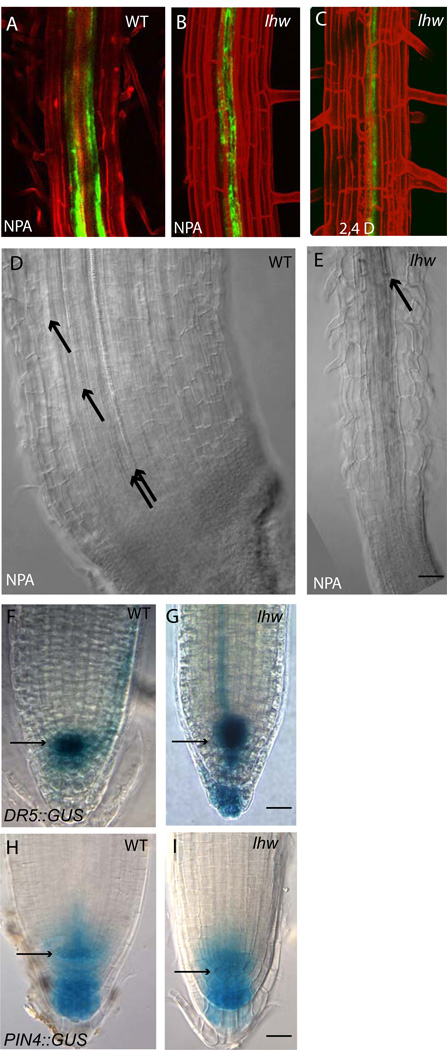
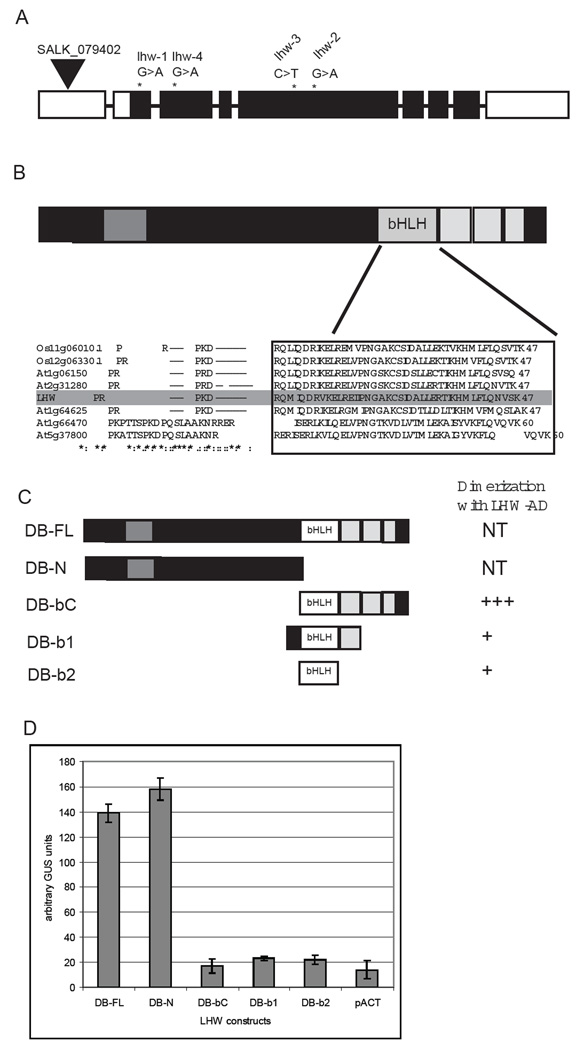
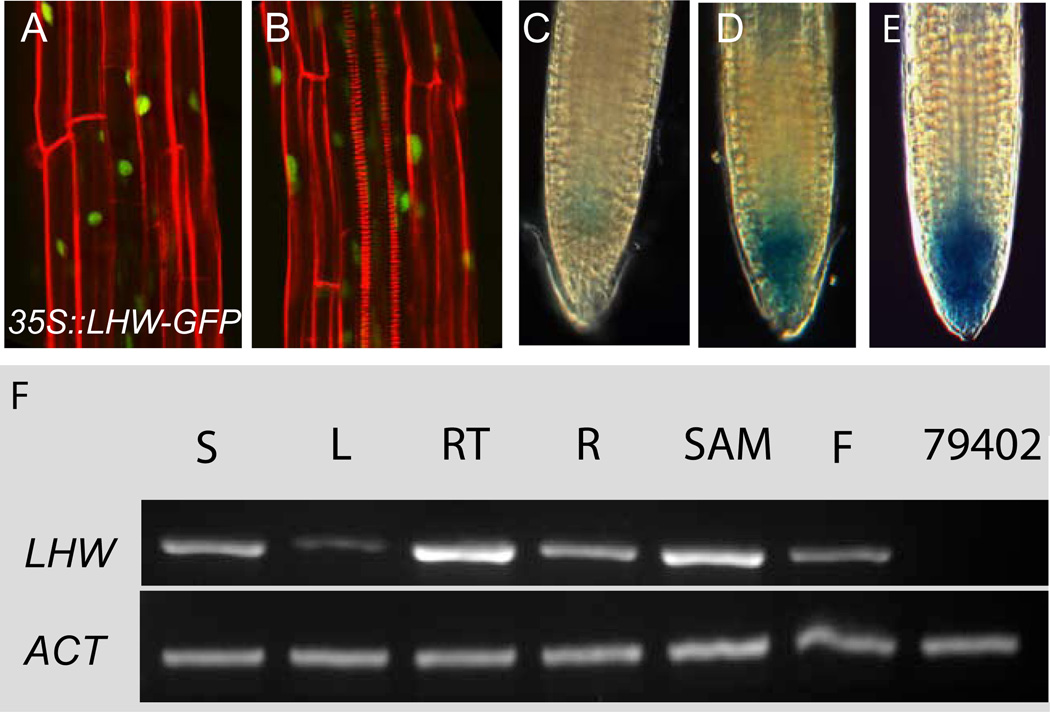
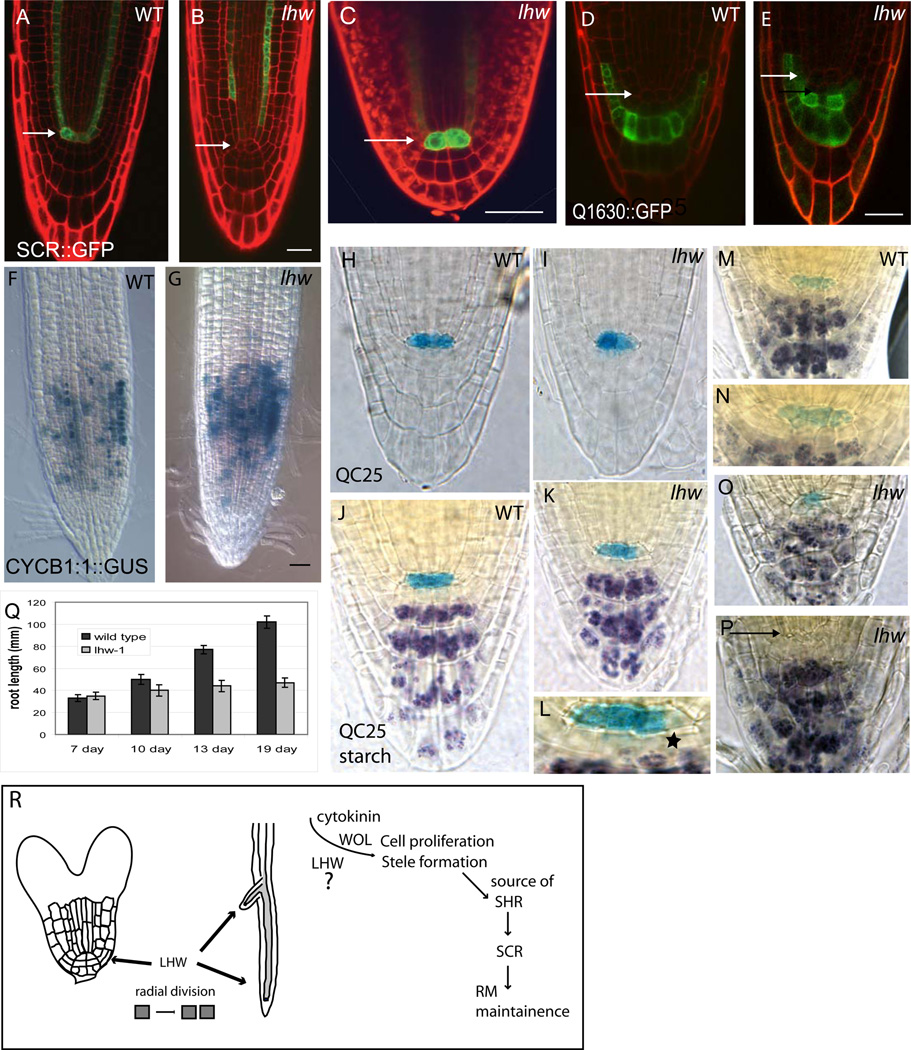
Complex organisms consist of a multitude of cell types arranged in a precise spatial relation to each other. Arabidopsis roots generally exhibit radial tissue organization; however, within a tissue layer, cells are not identical. Specific vascular cell types are arranged in diametrically opposed longitudinal files that maximize the distance between them and create a bilaterally symmetric (diarch) root. Mutations in the LONESOME HIGHWAY (LHW) gene eliminate bilateral symmetry and reduce the number of cells in the center of the root, resulting in roots with only single xylem and phloem poles. LHW does not appear to be required for the creation of any specific cell type, but coordinately controls the number of all vascular cell types by regulating the size of the pool of cells from which they arise. We cloned LHW and found that it encodes a protein with weak sequence similarity to basic helix-loop-helix (bHLH)-domain proteins. LHW is a transcriptional activator in vitro. In plants, LHW is nuclear-localized and is expressed in the root meristems, where we hypothesize it acts independently of other known root-patterning genes to promote the production of stele cells, but might also indirectly feed into established regulatory networks for the maintenance of the root meristem.
Figures
References
-
- Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, Galinha C, Nussaume L, Noh YS, Amasino R, Scheres B. The PLETHORA Genes Mediate Patterning of the Arabidopsis Root Stem Cell Niche. Cell. 2004;119:109–120. - PubMed
-
- Birnbaum K, Jung JW, Wang JY, Lambert GM, Hirst JA, Galbraith DW, Benfey PN. Cell Type-Specific Expression Profiling in Plants Via Cell Sorting of Protoplasts from Fluorescent Reporter Lines. Nat. Methods. 2005;2:615–619. - PubMed
-
- Birnbaum K, Shasha DE, Wang JY, Jung JW, Lambert GM, Galbraith DW, Benfey PN. A Gene Expression Map of the Arabidopsis Root. Science. 2003;302:1956–1960. - PubMed
-
- Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I, Friml J, Heidstra R, Aida M, Palme K, Scheres B. The PIN Auxin Efflux Facilitator Network Controls Growth and Patterning in Arabidopsis Roots. Nature. 2005;433:39–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
