

Enhanced at puberty 1 (EAP1) is a new transcriptional regulator of the female neuroendocrine reproductive axis
- PMID: 17627301
- PMCID: PMC1906733
- DOI: 10.1172/JCI31752
Enhanced at puberty 1 (EAP1) is a new transcriptional regulator of the female neuroendocrine reproductive axis
Abstract
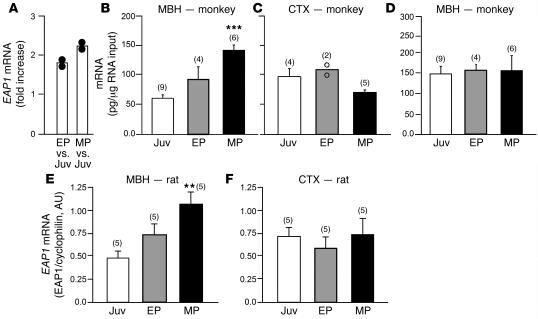
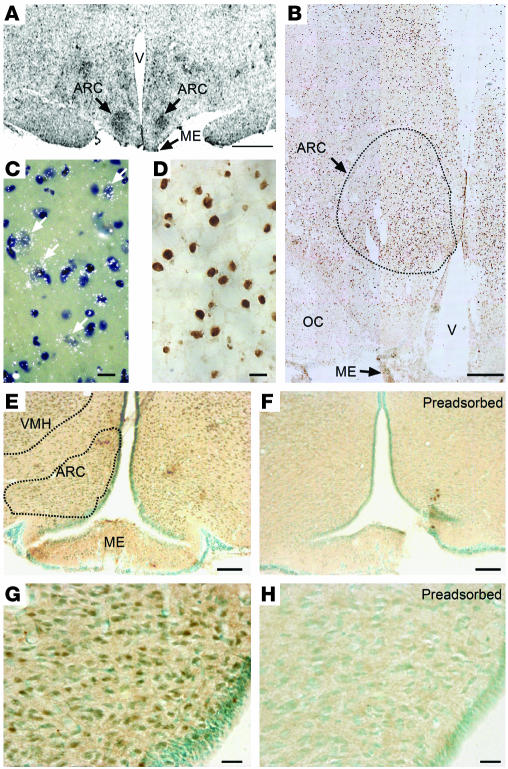
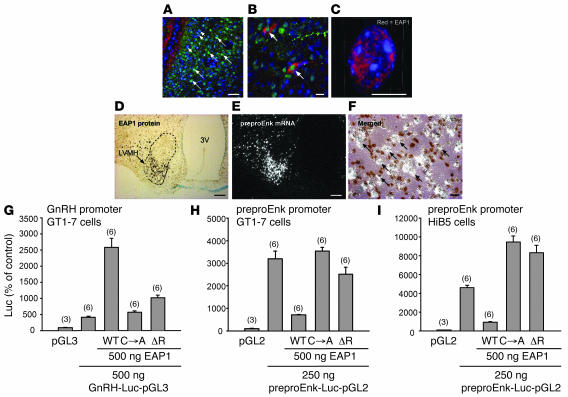
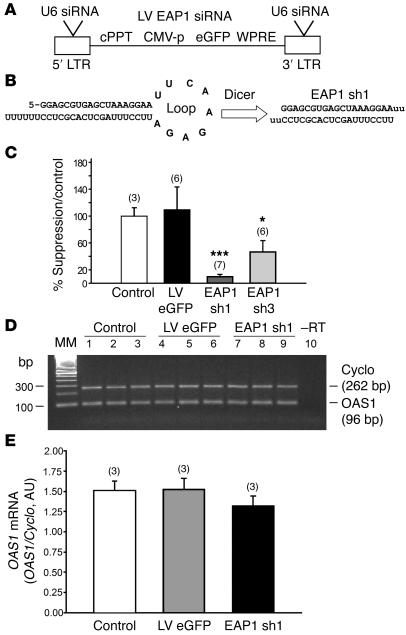
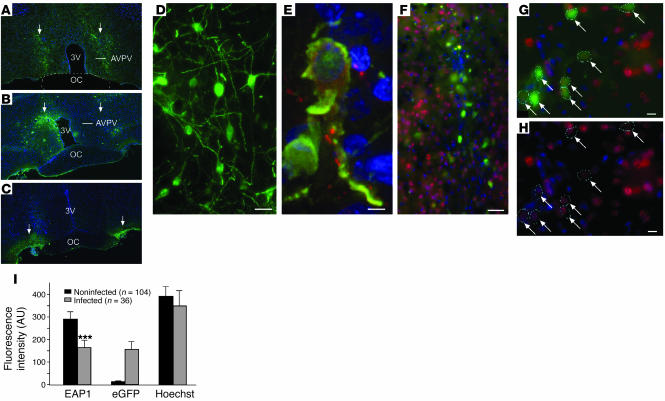
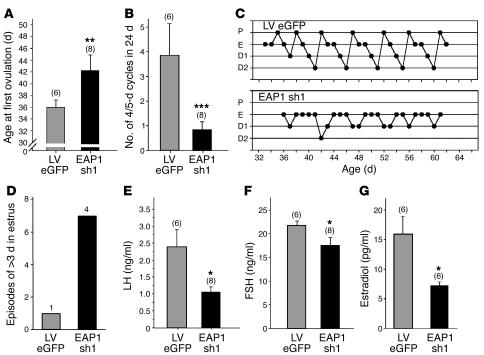
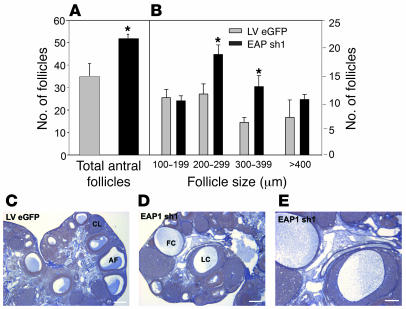
The initiation of mammalian puberty and the maintenance of female reproductive cycles are events controlled by hypothalamic neurons that secrete the decapeptide gonadotropin-releasing hormone (GnRH). GnRH secretion is, in turn, controlled by changes in neuronal and glial inputs to GnRH-producing neurons. The hierarchical control of the process is unknown, but it requires coordinated regulation of these cell-cell interactions. Here we report the functional characterization of a gene (termed enhanced at puberty 1 [EAP1]) that appears to act as an upstream transcriptional regulator of neuronal networks controlling female reproductive function. EAP1 expression increased selectively at puberty in both the nonhuman primate and rodent hypothalamus. EAP1 encoded a nuclear protein expressed in neurons involved in the inhibitory and facilitatory control of reproduction. EAP1 transactivated genes required for reproductive function, such as GNRH1, and repressed inhibitory genes, such as preproenkephalin. It contained a RING finger domain of the C3HC4 subclass required for this dual transcriptional activity. Inhibition of EAP1 expression, targeted to the rodent hypothalamus via lentivirus-mediated delivery of EAP1 siRNAs, delayed puberty, disrupted estrous cyclicity, and resulted in ovarian abnormalities. These results suggest that EAP1 is a transcriptional regulator that, acting within the neuroendocrine brain, contributes to controlling female reproductive function.
Figures
Similar articles
-
Transcription of the human EAP1 gene is regulated by upstream components of a puberty-controlling Tumor Suppressor Gene network.Mol Cell Endocrinol. 2012 Apr 4;351(2):184-98. doi: 10.1016/j.mce.2011.12.004. Epub 2011 Dec 19. Mol Cell Endocrinol. 2012. PMID: 22209758 Free PMC article.
-
Hypothalamic EAP1 (enhanced at puberty 1) is required for menstrual cyclicity in nonhuman primates.Endocrinology. 2012 Jan;153(1):350-61. doi: 10.1210/en.2011-1541. Epub 2011 Nov 29. Endocrinology. 2012. PMID: 22128022 Free PMC article.
-
Expression of EAP1 and CUX1 in the hypothalamus of female rats and relationship with KISS1 and GnRH.Endocr J. 2016 Aug 31;63(8):681-90. doi: 10.1507/endocrj.EJ16-0123. Epub 2016 May 31. Endocr J. 2016. PMID: 27250217
-
Glia-to-neuron signaling and the neuroendocrine control of female puberty.Recent Prog Horm Res. 2000;55:197-223; discussion 223-4. Recent Prog Horm Res. 2000. PMID: 11036938 Review.
-
Neuroendocrine mechanisms controlling female puberty: new approaches, new concepts.Int J Androl. 2006 Feb;29(1):256-63; discussion 286-90. doi: 10.1111/j.1365-2605.2005.00619.x. Int J Androl. 2006. PMID: 16466547 Review.
Cited by
-
A novel IRF2BPL truncating variant is associated with endolysosomal storage.Mol Biol Rep. 2020 Jan;47(1):711-714. doi: 10.1007/s11033-019-05109-7. Epub 2019 Oct 3. Mol Biol Rep. 2020. PMID: 31583567
-
Genetic and genomic characterization of vulva size traits in Yorkshire and Landrace gilts.BMC Genet. 2020 Mar 12;21(1):28. doi: 10.1186/s12863-020-0834-9. BMC Genet. 2020. PMID: 32164558 Free PMC article.
-
Changes in hypothalamic expression of the Lin28/let-7 system and related microRNAs during postnatal maturation and after experimental manipulations of puberty.Endocrinology. 2013 Feb;154(2):942-55. doi: 10.1210/en.2012-2006. Epub 2013 Jan 4. Endocrinology. 2013. PMID: 23291449 Free PMC article.
-
Genes underlying delayed puberty.Mol Cell Endocrinol. 2018 Nov 15;476:119-128. doi: 10.1016/j.mce.2018.05.001. Epub 2018 May 4. Mol Cell Endocrinol. 2018. PMID: 29730183 Free PMC article. Review.
-
Effects of 4-vinylcyclohexene diepoxide on peripubertal and adult Sprague-Dawley rats: ovarian, clinical, and pathologic outcomes.Comp Med. 2009 Feb;59(1):46-59. Comp Med. 2009. PMID: 19295054 Free PMC article.
References
-
- Plant, T.M., and Witchel, S.F. 2006. Puberty in nonhuman primates and humans. In The physiology of reproduction. 3rd edition. J.D. Neill, editor. Academic Press/Elsevier. San Diego, California, USA. 2177–2230.
-
- Ojeda, S.R., and Skinner, M.K. 2006. Puberty in the rat. In The physiology of reproduction. 3rd edition. J.D. Neill, editor. Academic Press/Elsevier. San Diego, California, USA. 2061–2126.
-
- Kordon, C., Drouva, S.V., Martínez de la Escalera, G., and Weiner, R.I. 1994. Role of classic and peptide neuromediators in the neuroendocrine regulation of luteinizing hormone and prolactin. In The physiology of reproduction. 2nd edition. Volume 1. E. Knobil and J.D. Neill, editors. Raven Press. New York, New York, USA. 1621–1681.
-
- Ojeda, S.R., and Terasawa, E. 2002. Neuroendocrine regulation of puberty. In Hormones, brain and behavior. Volume 4. D. Pfaff et al., editors. Elsevier. New York, New York, USA. 589–659.
-
- Ojeda S.R., et al. Glia-to-neuron signaling and the neuroendocrine control of female puberty. Ann. Med. 2003;35:244–255. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
