

Histamine excites neurones in the human submucous plexus through activation of H1, H2, H3 and H4 receptors
- PMID: 17627982
- PMCID: PMC2277025
- DOI: 10.1113/jphysiol.2007.139352
Histamine excites neurones in the human submucous plexus through activation of H1, H2, H3 and H4 receptors
Abstract
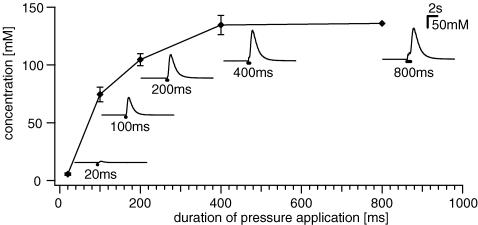
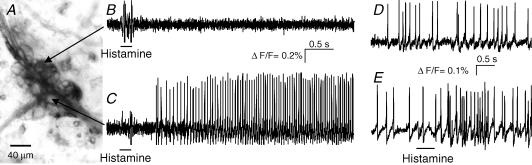
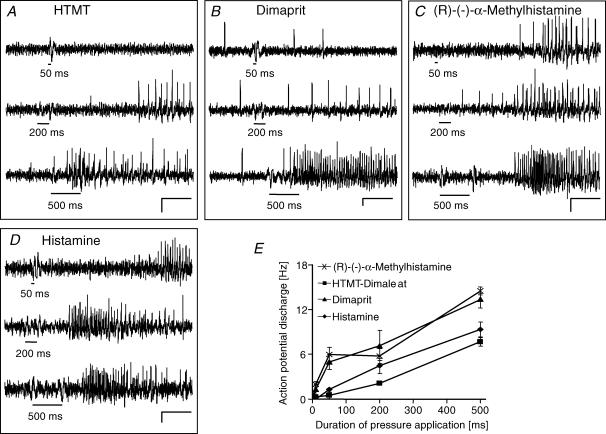
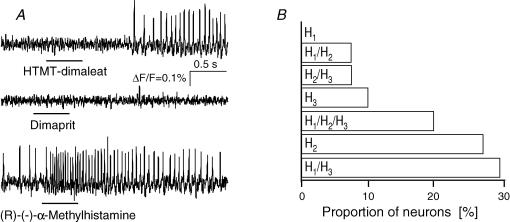
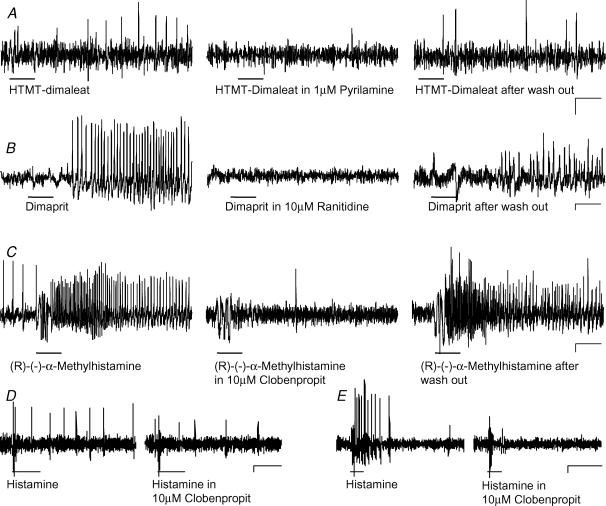
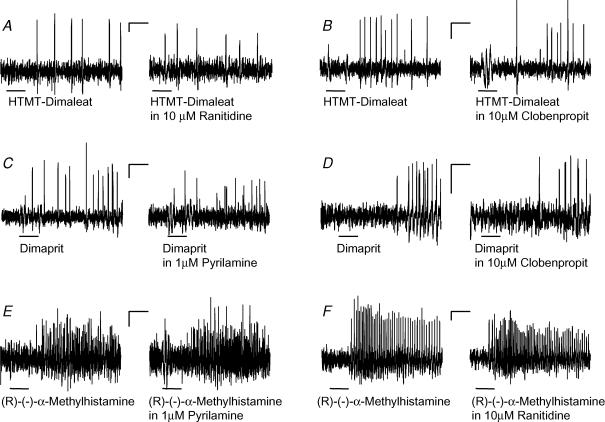
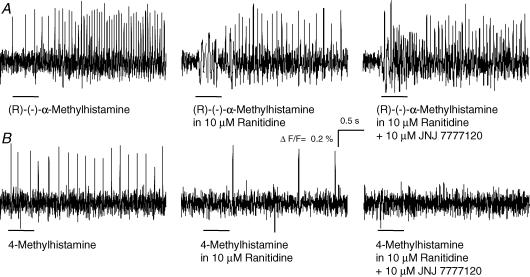
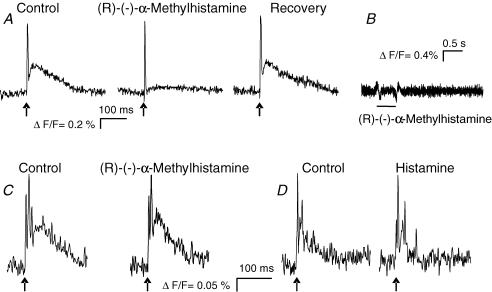
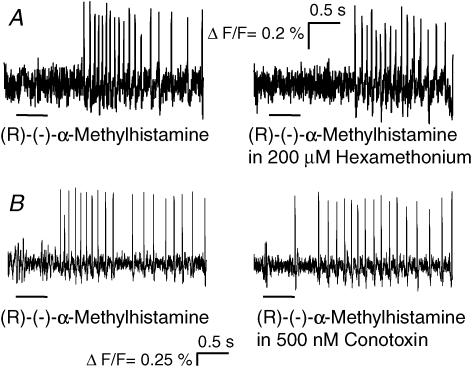
Histamine is a major mast cell mediator of immunoneural signalling in the gut and mast cells play a role in the pathophysiology of functional and inflammatory bowel diseases. Histamine receptors are therefore promising drug targets to treat gut disorders. We aimed to study the so far unknown effect of histamine on neural activity in the human enteric nervous system (ENS) and to identify the pharmacology of histamine response. We used fast imaging techniques in combination with the potentiometric dye di-8-ANEPPS to monitor directly membrane potential changes and thereby neuronal excitability in the human submucous plexus from surgical specimens of 110 patients (2137 neurones, 273 ganglia). Local microejection of histamine resulted in action potential discharge in 37% of neurones. This excitatory effect was mimicked by the H(1) agonist HTMT-dimaleat, H(2) agonist dimaprit, H(3) agonist (R)-(-)-alpha-methylhistamine and H(4) agonist 4-methylhistamine. The excitatory actions of the agonists were specifically and selectively blocked by the H(1), H(2), H(3) or H(4) receptor antagonists pyrilamine, ranitidine, clobenpropit or J1-[(5-chloro-1H-indol-2-yl)carbonyl]-4-methylpiperazine (JNJ 7777120), respectively. Clobenproprit reduced the excitatory response to histamine. Unlike in the guinea-pig ENS (R)-(-)-alpha-methylhistamine had no presynaptic actions in human submucous plexus. Application of agonists revealed receptor clustering which was as follows: 29% H(1)/H(3), 27% H(2), 20% H(1)/H(2)/H(3), 10% H(3), 7% H(1)/H(2) and 7% H(2)/H(3). Histamine excites human enteric neurones and this effect involves all four histamine receptors; most striking was the identification of an excitatory H(3) mediated component and the discovery of H(4) mediated neuronal excitation. These data may form the basis of identification of new targets to treat inflammatory and functional gut disorders.
Figures
References
-
- Bakker RA. Histamine H3-receptor isoforms. Inflamm Res. 2004;53:509–516. - PubMed
-
- Barbara G, Stanghellini V, de Giorgio R, Corinaldesi R. Functional gastrointestinal disorders and mast cells: implications for therapy. Neurogastroenterol Motil. 2006;18:6–17. - PubMed
-
- Barbara G, Stanghellini V, De Giorgio R, Cremon C, Cottrell GS, Santini D, Pasquinelli G, Morselli-Labate AM, Grady EF, Bunnett NW, Collins SM, Corinaldesi R. Activated mast cells in proximity to colonic nerves correlate with abdominal pain in irritable bowel syndrome. Gastroenterology. 2004;126:693–702. - PubMed
-
- Bischoff S, Crowe SE. Gastrointestinal food allergy: new insights into pathophysiology and clinical perspectives. Gastroenterology. 2005;128:1089–1113. - PubMed
-
- Celanire S, Wijtmans M, Talaga P, Leurs R, de Esch IJ. Keynote review: Histamine H3 receptor antagonists reach out for the clinic. Drug Discov Today. 2005;10:1613–1627. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources