

Predictive oncology: a review of multidisciplinary, multiscale in silico modeling linking phenotype, morphology and growth
- PMID: 17629503
- PMCID: PMC2245890
- DOI: 10.1016/j.neuroimage.2007.05.043
Predictive oncology: a review of multidisciplinary, multiscale in silico modeling linking phenotype, morphology and growth
Abstract
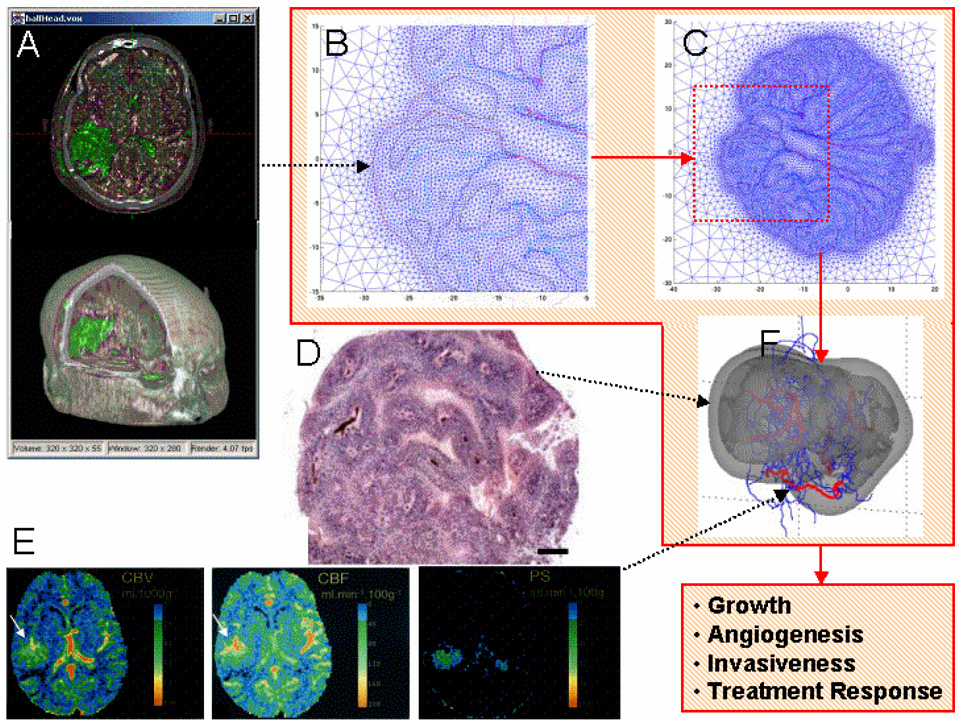
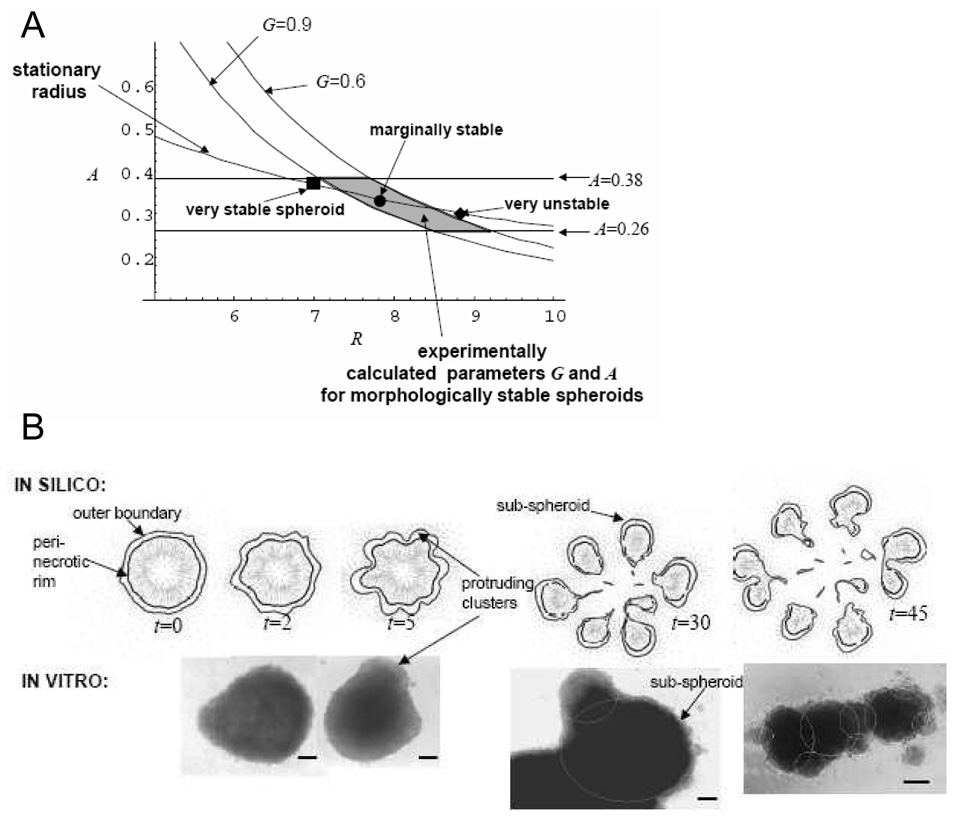
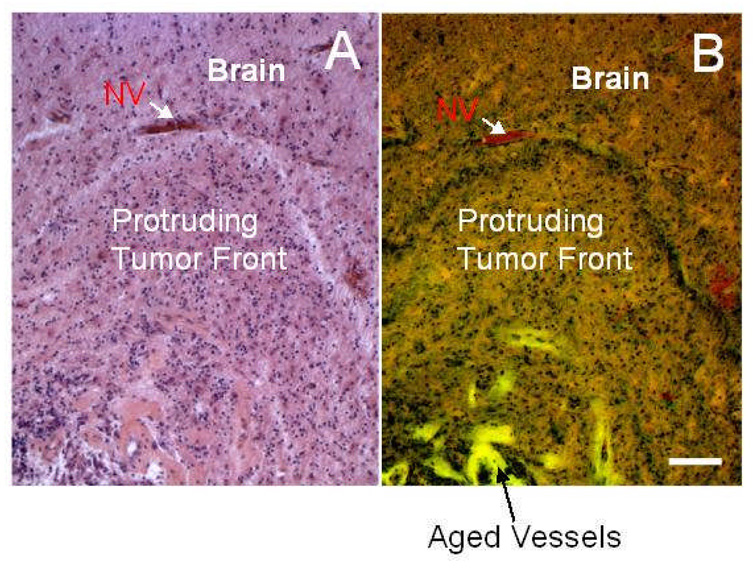
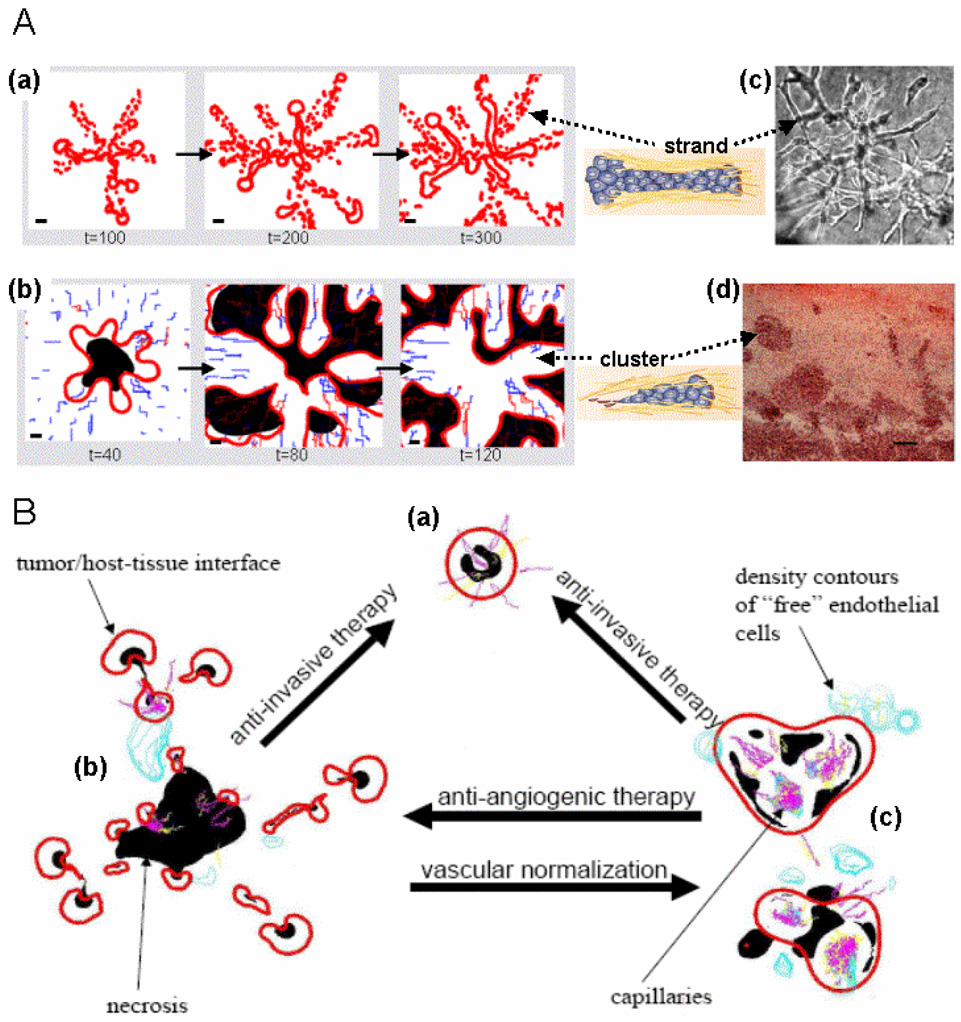
Empirical evidence and theoretical studies suggest that the phenotype, i.e., cellular- and molecular-scale dynamics, including proliferation rate and adhesiveness due to microenvironmental factors and gene expression that govern tumor growth and invasiveness, also determine gross tumor-scale morphology. It has been difficult to quantify the relative effect of these links on disease progression and prognosis using conventional clinical and experimental methods and observables. As a result, successful individualized treatment of highly malignant and invasive cancers, such as glioblastoma, via surgical resection and chemotherapy cannot be offered and outcomes are generally poor. What is needed is a deterministic, quantifiable method to enable understanding of the connections between phenotype and tumor morphology. Here, we critically assess advantages and disadvantages of recent computational modeling efforts (e.g., continuum, discrete, and cellular automata models) that have pursued this understanding. Based on this assessment, we review a multiscale, i.e., from the molecular to the gross tumor scale, mathematical and computational "first-principle" approach based on mass conservation and other physical laws, such as employed in reaction-diffusion systems. Model variables describe known characteristics of tumor behavior, and parameters and functional relationships across scales are informed from in vitro, in vivo and ex vivo biology. We review the feasibility of this methodology that, once coupled to tumor imaging and tumor biopsy or cell culture data, should enable prediction of tumor growth and therapy outcome through quantification of the relation between the underlying dynamics and morphological characteristics. In particular, morphologic stability analysis of this mathematical model reveals that tumor cell patterning at the tumor-host interface is regulated by cell proliferation, adhesion and other phenotypic characteristics: histopathology information of tumor boundary can be inputted to the mathematical model and used as a phenotype-diagnostic tool to predict collective and individual tumor cell invasion of surrounding tissue. This approach further provides a means to deterministically test effects of novel and hypothetical therapy strategies on tumor behavior.
Figures




References
-
- Adam J. General aspects of modeling tumor growth and the immune response. In: Adam J, Bellomo N, editors. A survey of models on tumor immune systems dynamics. Boston: Birkhauser; 1996. pp. 15–87.
-
- Ambrosi D, Guana F. Stress-modulated growth. Math. Meth. Solids. 2006 in press.
-
- Ambrosi D, Preziosi L. On the closure of mass balance models for tumor growth. Math. Mod. Meth. Appl. Sci. 2002;12:737–754.
-
- Anderson ARA, Chaplain MAJ. Continuous and discrete models of tumour-induced angiogenesis. Bull. Math. Biol. 1998;60:857–899. - PubMed
-
- Anderson ARA, Chaplain MAJ, Newman EL, Steele RJC, Thompson AM. Mathematical modeling of tumour invasion and metastasis. J. Theor. Med. 2000;2:129–154.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
