

Simulation of networks of spiking neurons: a review of tools and strategies
- PMID: 17629781
- PMCID: PMC2638500
- DOI: 10.1007/s10827-007-0038-6
Simulation of networks of spiking neurons: a review of tools and strategies
Abstract
We review different aspects of the simulation of spiking neural networks. We start by reviewing the different types of simulation strategies and algorithms that are currently implemented. We next review the precision of those simulation strategies, in particular in cases where plasticity depends on the exact timing of the spikes. We overview different simulators and simulation environments presently available (restricted to those freely available, open source and documented). For each simulation tool, its advantages and pitfalls are reviewed, with an aim to allow the reader to identify which simulator is appropriate for a given task. Finally, we provide a series of benchmark simulations of different types of networks of spiking neurons, including Hodgkin-Huxley type, integrate-and-fire models, interacting with current-based or conductance-based synapses, using clock-driven or event-driven integration strategies. The same set of models are implemented on the different simulators, and the codes are made available. The ultimate goal of this review is to provide a resource to facilitate identifying the appropriate integration strategy and simulation tool to use for a given modeling problem related to spiking neural networks.
Figures



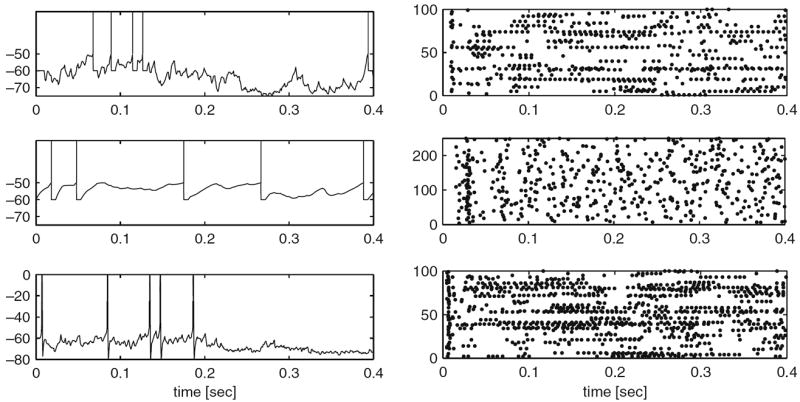
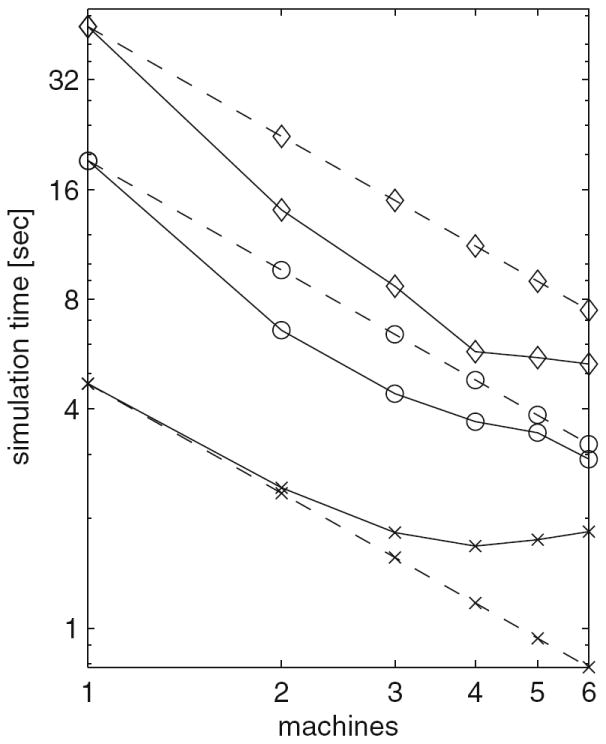
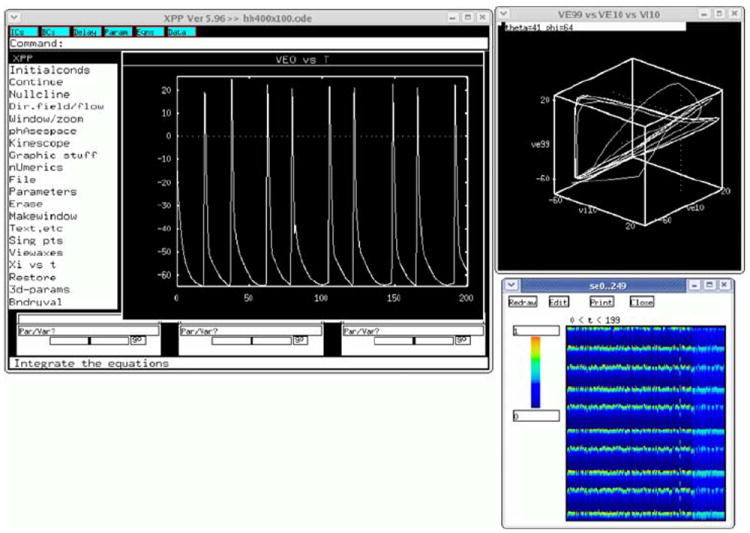
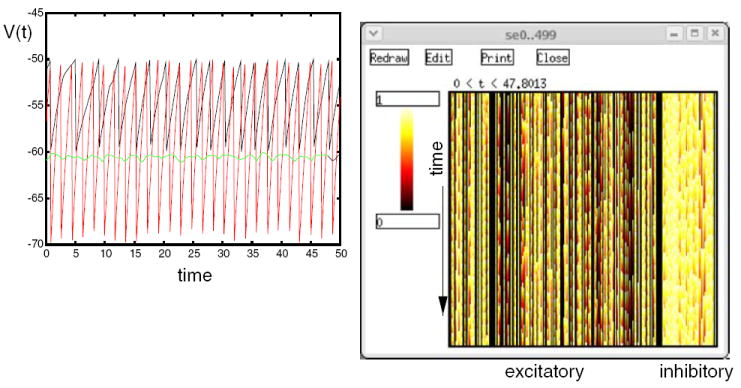
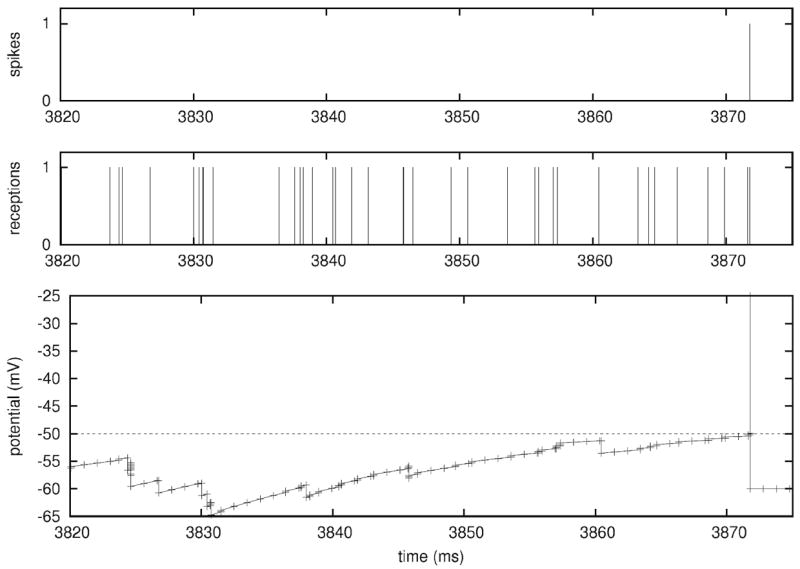

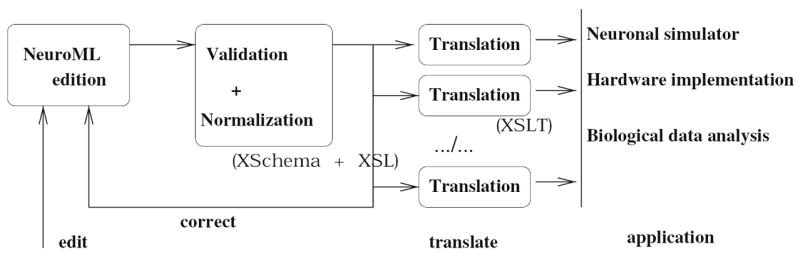

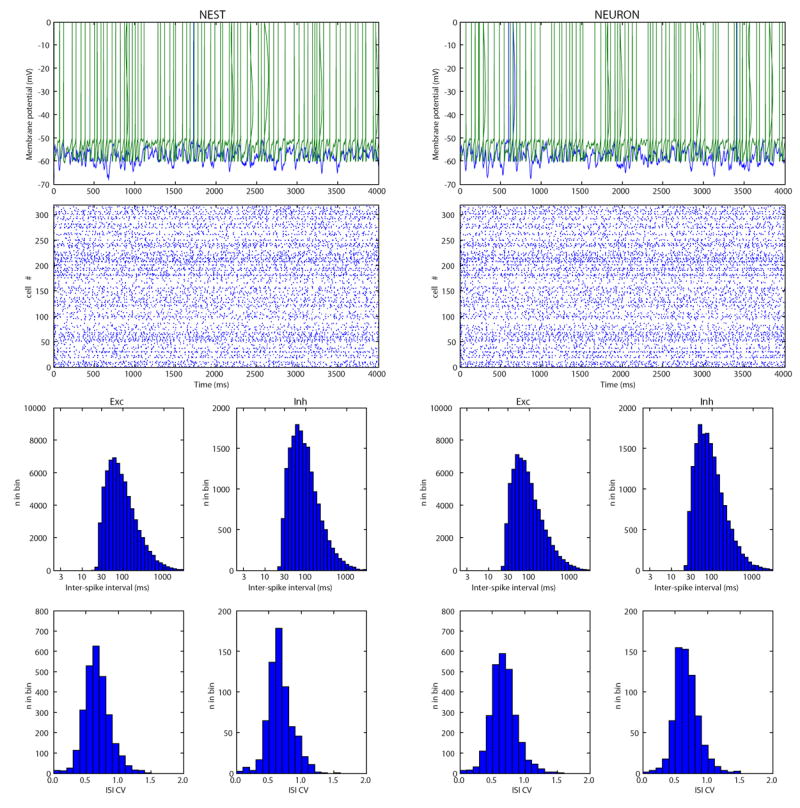
References
-
- Abbott LF, Nelson SB. Synaptic plasticity: taming the beast. Nature Neuroscience. 2000;3(Suppl):1178–1283. - PubMed
-
- Arnold L. Stochastic differential equations: Theory and applications. New York: J Wiley and Sons; 1974.
-
- Azouz R. Dynamic spatiotemporal synaptic integration in cortical neurons: neuronal gain, revisited. Journal of Neurophysiology. 2005;94:2785–2796. - PubMed
-
- Badoual M, Rudolph M, Piwkowska Z, Destexhe A, Bal T. High discharge variability in neurons driven by current noise. Neurocomputing. 2005;65:493–498.
-
- Bailey J, Hammerstrom D. International Conference on Neural Networks (ICNN 88, IEEE) San Diego: 1988. Why VLSI implementations of associative VLCNs require connection multiplexing; pp. 173–180.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
