

A unique beta1,3-galactosyltransferase is indispensable for the biosynthesis of N-glycans containing Lewis a structures in Arabidopsis thaliana
- PMID: 17630273
- PMCID: PMC1955701
- DOI: 10.1105/tpc.107.052985
A unique beta1,3-galactosyltransferase is indispensable for the biosynthesis of N-glycans containing Lewis a structures in Arabidopsis thaliana
Abstract
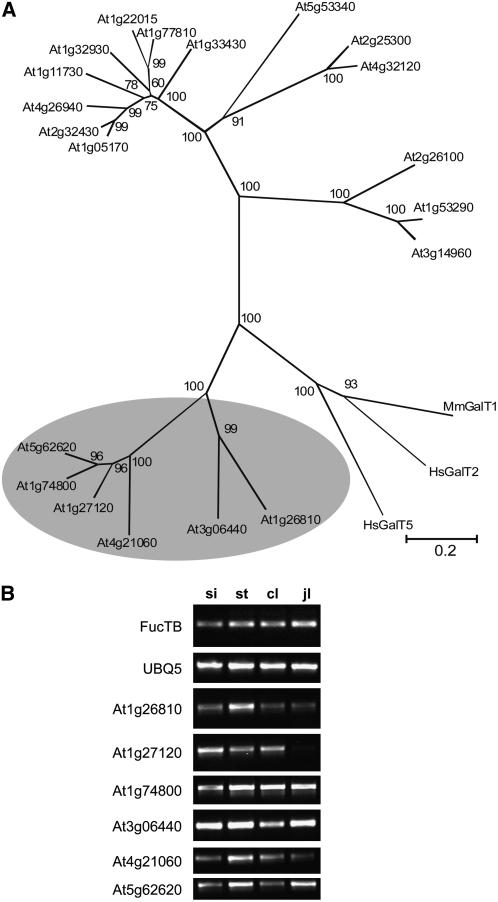
In plants, the only known outer-chain elongation of complex N-glycans is the formation of Lewis a [Fuc alpha1-4(Gal beta1-3)GlcNAc-R] structures. This process involves the sequential attachment of beta1,3-galactose and alpha1,4-fucose residues by beta1,3-galactosyltransferase and alpha1,4-fucosyltransferase. However, the exact mechanism underlying the formation of Lewis a epitopes in plants is poorly understood, largely because one of the involved enzymes, beta1,3-galactosyltransferase, has not yet been identified and characterized. Here, we report the identification of an Arabidopsis thaliana beta1,3-galactosyltransferase involved in the biosynthesis of the Lewis a epitope using an expression cloning strategy. Overexpression of various candidates led to the identification of a single gene (named GALACTOSYLTRANSFERASE1 [GALT1]) that increased the originally very low Lewis a epitope levels in planta. Recombinant GALT1 protein produced in insect cells was capable of transferring beta1,3-linked galactose residues to various N-glycan acceptor substrates, and subsequent treatment of the reaction products with alpha1,4-fucosyltransferase resulted in the generation of Lewis a structures. Furthermore, transgenic Arabidopsis plants lacking a functional GALT1 mRNA did not show any detectable amounts of Lewis a epitopes on endogenous glycoproteins. Taken together, our results demonstrate that GALT1 is both sufficient and essential for the addition of beta1,3-linked galactose residues to N-glycans and thus is required for the biosynthesis of Lewis a structures in Arabidopsis. Moreover, cell biological characterization of a transiently expressed GALT1-fluorescent protein fusion using confocal laser scanning microscopy revealed the exclusive location of GALT1 within the Golgi apparatus, which is in good agreement with the proposed physiological action of the enzyme.
Figures










Similar articles
-
Plant protein glycosylation.Glycobiology. 2016 Sep;26(9):926-939. doi: 10.1093/glycob/cww023. Epub 2016 Feb 23. Glycobiology. 2016. PMID: 26911286 Free PMC article. Review.
-
Galactosyltransferases from Arabidopsis thaliana in the biosynthesis of type II arabinogalactan: molecular interaction enhances enzyme activity.BMC Plant Biol. 2014 Apr 3;14:90. doi: 10.1186/1471-2229-14-90. BMC Plant Biol. 2014. PMID: 24693939 Free PMC article.
-
Identification of β1,3-galactosyltransferases responsible for biosynthesis of insect complex-type N-glycans containing a T-antigen unit in the honeybee.Glycoconj J. 2015 May;32(3-4):141-51. doi: 10.1007/s10719-015-9585-7. Epub 2015 May 1. Glycoconj J. 2015. PMID: 25931033
-
Arabidopsis β1,2-xylosyltransferase: substrate specificity and participation in the plant-specific N-glycosylation pathway.J Biosci Bioeng. 2012 Jan;113(1):48-54. doi: 10.1016/j.jbiosc.2011.09.011. Epub 2011 Oct 24. J Biosci Bioeng. 2012. PMID: 22024534
-
The galactosyltransferase family.Cell Mol Life Sci. 2002 Jul;59(7):1081-95. doi: 10.1007/s00018-002-8489-4. Cell Mol Life Sci. 2002. PMID: 12222957 Free PMC article. Review.
Cited by
-
Plant protein glycosylation.Glycobiology. 2016 Sep;26(9):926-939. doi: 10.1093/glycob/cww023. Epub 2016 Feb 23. Glycobiology. 2016. PMID: 26911286 Free PMC article. Review.
-
Arabinogalactan-proteins: key regulators at the cell surface?Plant Physiol. 2010 Jun;153(2):403-19. doi: 10.1104/pp.110.156000. Epub 2010 Apr 13. Plant Physiol. 2010. PMID: 20388666 Free PMC article. Review. No abstract available.
-
N-glycan Remodeling Using Mannosidase Inhibitors to Increase High-mannose Glycans on Acid α-Glucosidase in Transgenic Rice Cell Cultures.Sci Rep. 2018 Oct 31;8(1):16130. doi: 10.1038/s41598-018-34438-z. Sci Rep. 2018. PMID: 30382146 Free PMC article.
-
Improved recombinant protein production in Arabidopsis thaliana.Plant Signal Behav. 2018;13(6):e1486149. doi: 10.1080/15592324.2018.1486149. Epub 2018 Jun 22. Plant Signal Behav. 2018. PMID: 29932798 Free PMC article.
-
Stable Protein Sialylation in Physcomitrella.Front Plant Sci. 2020 Dec 18;11:610032. doi: 10.3389/fpls.2020.610032. eCollection 2020. Front Plant Sci. 2020. PMID: 33391325 Free PMC article.
References
-
- Ackerman, S., Liu, L., Kwatia, M., Savage, M., Leonidas, D., Swaminathan, G., and Acharya, K. (2002). Charcot-Leyden crystal protein (galectin-10) is not a dual function galectin with lysophospholipase activity but binds a lysophospholipase inhibitor in a novel structural fashion. J. Biol. Chem. 277 14859–14868. - PubMed
-
- Altmann, F., Schweiszer, S., and Weber, C. (1995). Kinetic comparison of peptide: N-glycosidases F and A reveals several differences in substrate specificity. Glycoconj. J. 12 84–93. - PubMed
-
- Amado, M., Almeida, R., Carneiro, F., Levery, S., Holmes, E., Nomoto, M., Hollingsworth, M., Hassan, H., Schwientek, T., Nielsen, P., Bennett, E., and Clausen, H. (1998). A family of human beta3-galactosyltransferases. Characterization of four members of a UDP-galactose:beta-N-acetyl-glucosamine/beta-nacetyl-galactosamine beta-1,3-galactosyltransferase family. J. Biol. Chem. 273 12770–12778. - PubMed
-
- Bakker, H., Schijlen, E., de Vries, T., Schiphorst, W., Jordi, W., Lommen, A., Bosch, D., and van Die, I. (2001. a). Plant members of the alpha1-3/4-fucosyltransferase gene family encode an alpha1-4-fucosyltransferase, potentially involved in Lewis(a) biosynthesis, and two core alpha1-3-fucosyltransferases. FEBS Lett. 507 307–312. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
