

Nuclear lamin A/C deficiency induces defects in cell mechanics, polarization, and migration
- PMID: 17631533
- PMCID: PMC1965451
- DOI: 10.1529/biophysj.106.102426
Nuclear lamin A/C deficiency induces defects in cell mechanics, polarization, and migration
Abstract
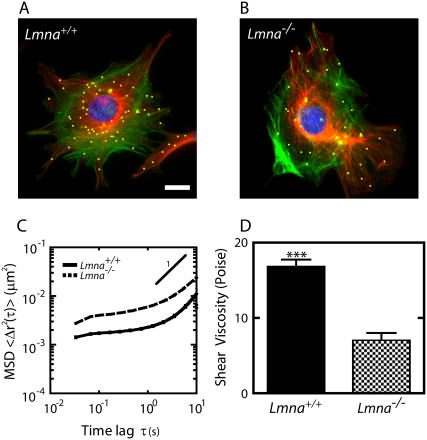
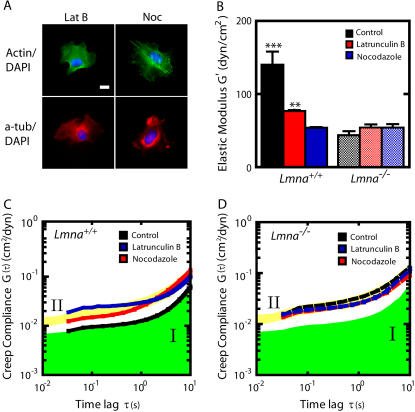
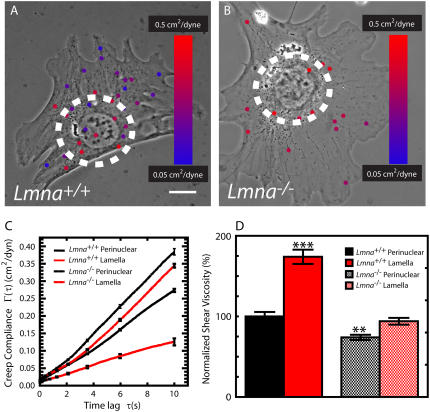
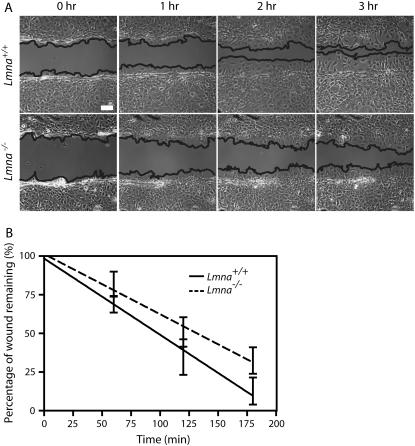
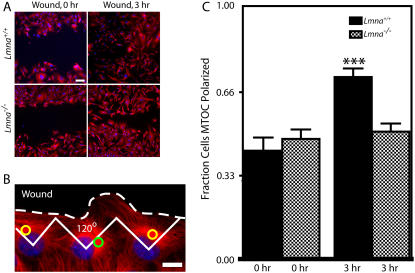
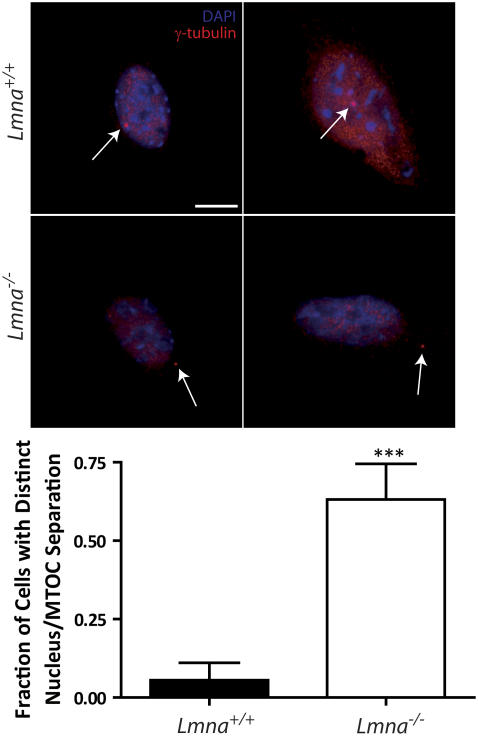
Lamin A/C is a major constituent of the nuclear lamina, a thin filamentous protein layer that lies beneath the nuclear envelope. Here we show that lamin A/C deficiency in mouse embryonic fibroblasts (Lmna(-/-) MEFs) diminishes the ability of these cells to polarize at the edge of a wound and significantly reduces cell migration speed into the wound. Moreover, lamin A/C deficiency induces significant separation of the microtubule organizing center (MTOC) from the nuclear envelope. Investigations using ballistic intracellular nanorheology reveal that lamin A/C deficiency also dramatically affects the micromechanical properties of the cytoplasm. Both the elasticity (stretchiness) and the viscosity (propensity of a material to flow) of the cytoplasm in Lmna(-/-) MEFs are significantly reduced. Disassembly of either the actin filament or microtubule networks in Lmna(+/+) MEFs results in decrease of cytoplasmic elasticity and viscosity down to levels found in Lmna(-/-) MEFs. Together these results show that both the mechanical properties of the cytoskeleton and cytoskeleton-based processes, including cell motility, coupled MTOC and nucleus dynamics, and cell polarization, depend critically on the integrity of the nuclear lamina, which suggest the existence of a functional mechanical connection between the nucleus and the cytoskeleton. These results also suggest that cell polarization during cell migration requires tight mechanical coupling between MTOC and nucleus, which is mediated by lamin A/C.
Figures
References
-
- Hetzer, M. W., T. C. Walther, and I. W. Mattaj. 2005. Pushing the envelope: structure, function, and dynamics of the nuclear periphery. Annu. Rev. Cell Dev. Biol. 21:347–380. - PubMed
-
- Hutchison, C. J. 2002. Lamins: building blocks or regulators of gene expression? Nat. Rev. Mol. Cell Biol. 3:848–858. - PubMed
-
- Stuurman, N., S. Heins, and U. Aebi. 1998. Nuclear lamins: their structure, assembly, and interactions. J. Struct. Biol. 122:42–66. - PubMed
-
- Elbashir, S. M., J. Harborth, W. Lendeckel, A. Yalcin, K. Weber, and T. Tuschl. 2001. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 411:494–498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
