

Iron chelators ICL670 and 311 inhibit HIV-1 transcription
- PMID: 17631934
- PMCID: PMC2077891
- DOI: 10.1016/j.virol.2007.06.011
Iron chelators ICL670 and 311 inhibit HIV-1 transcription
Abstract
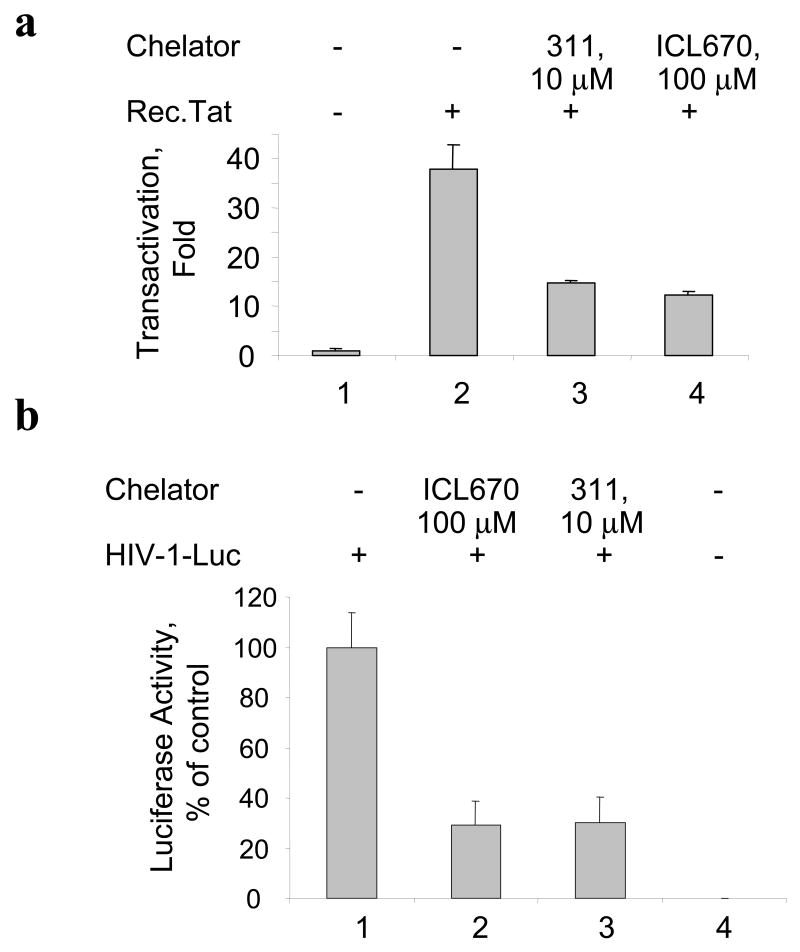
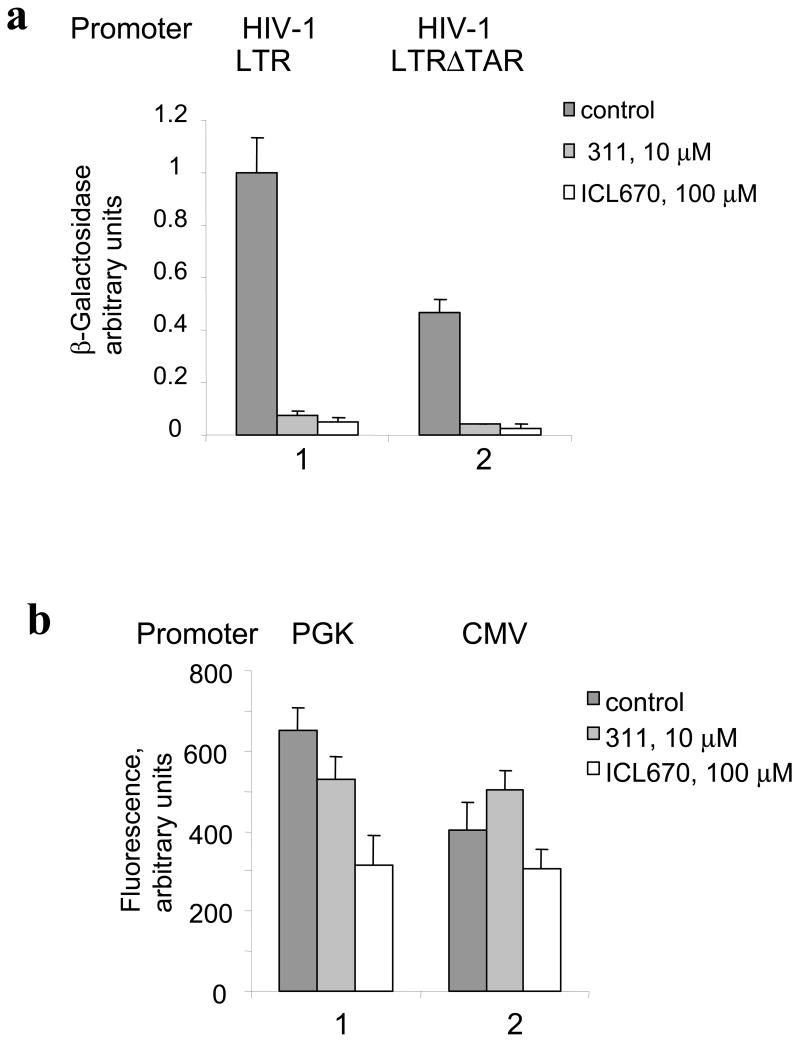
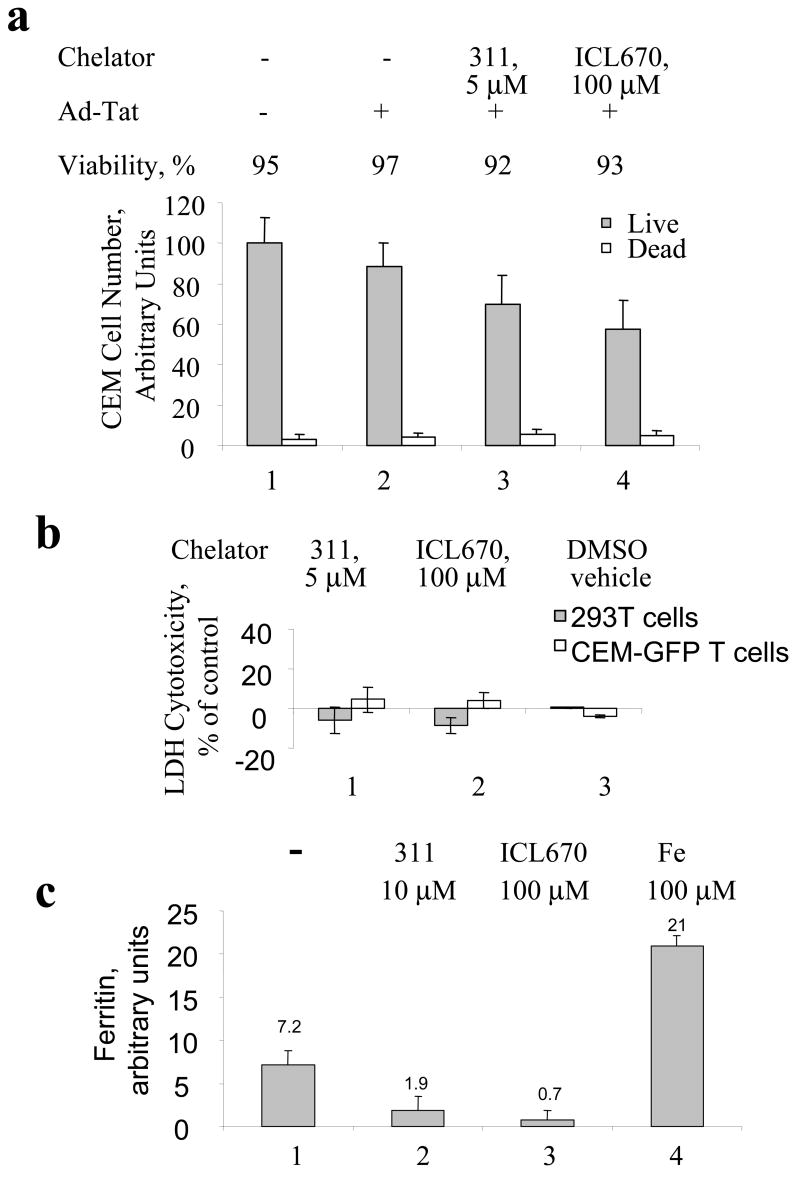
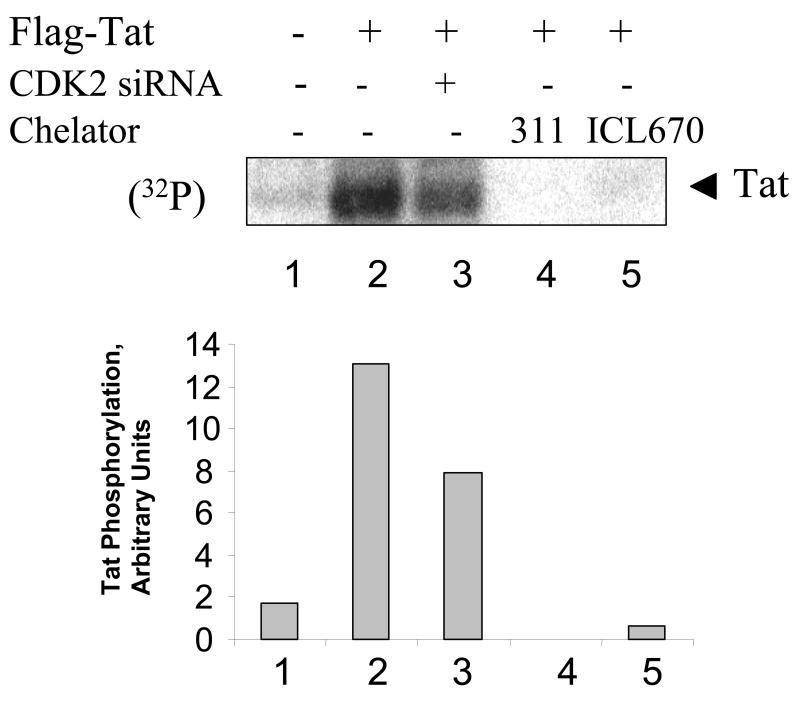
HIV-1 replication is induced by an excess of iron and iron chelation by desferrioxamine (DFO) inhibits viral replication by reducing proliferation of infected cells. Treatment of cells with DFO and 2-hydroxy-1-naphthylaldehyde isonicotinoyl hydrazone (311) inhibit expression of proteins that regulate cell-cycle progression, including cycle-dependent kinase 2 (CDK2). Our recent studies showed that CDK2 participates in HIV-1 transcription and viral replication suggesting that inhibition of CDK2 by iron chelators might also affect HIV-1 transcription. Here we evaluated the effect of a clinically approved orally effective iron chelator, 4-[3,5-bis-(hydroxyphenyl)-1,2,4-triazol-1-yl]-benzoic acid (ICL670) and 311 on HIV-1 transcription. Both ICL670 and 311 inhibited Tat-induced HIV-1 transcription in CEM-T cells, 293T and HeLa cells. Neither ICL670 nor 311 induced cytotoxicity at concentrations that inhibited HIV-1 transcription. The chelators decreased cellular activity of CDK2 and reduced HIV-1 Tat phosphorylation by CDK2. Neither ICL670A or 311 decreased CDK9 protein level but significantly reduced association of CDK9 with cyclin T1 and reduced phosphorylation of Ser-2 residues of RNA polymerase II C-terminal domain. In conclusion, our findings add to the evidence that iron chelators can inhibit HIV-1 transcription by deregulating CDK2 and CDK9. Further consideration should be given to the development of iron chelators for future anti-retroviral therapeutics.
Figures



References
-
- Agbottah E, de la Fuente C, Nekhai S, Barnett A, Gianella-Borradori A, Pumfery A, Kashanchi F. Antiviral activity of CYC202 in HIV-1 infected cells. J Biol Chem 2004 - PubMed
-
- Agbottah E, de La Fuente C, Nekhai S, Barnett A, Gianella-Borradori A, Pumfery A, Kashanchi F. Antiviral activity of CYC202 in HIV-1-infected cells. J Biol Chem. 2005;280(4):3029–42. - PubMed
-
- Ammosova T, Berro R, Kashanchi F, Nekhai S. RNA interference directed to CDK2 inhibits HIV-1 transcription. Virology. 2005a;341(2):171–8. - PubMed
-
- Ammosova T, Jerebtsova M, Beullens M, Lesage B, Jackson A, Kashanchi F, Southerland W, Gordeuk VR, Bollen M, Nekhai S. Nuclear targeting of protein phosphatase-1 by HIV-1 Tat protein. J Biol Chem. 2005b;280(43):36364–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
