

Unusual transformations in the biosynthesis of the antibiotic phosphinothricin tripeptide
- PMID: 17632514
- PMCID: PMC4313788
- DOI: 10.1038/nchembio.2007.9
Unusual transformations in the biosynthesis of the antibiotic phosphinothricin tripeptide
Abstract
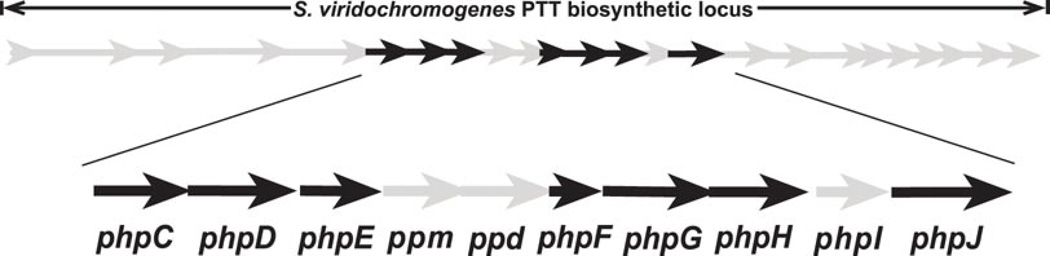
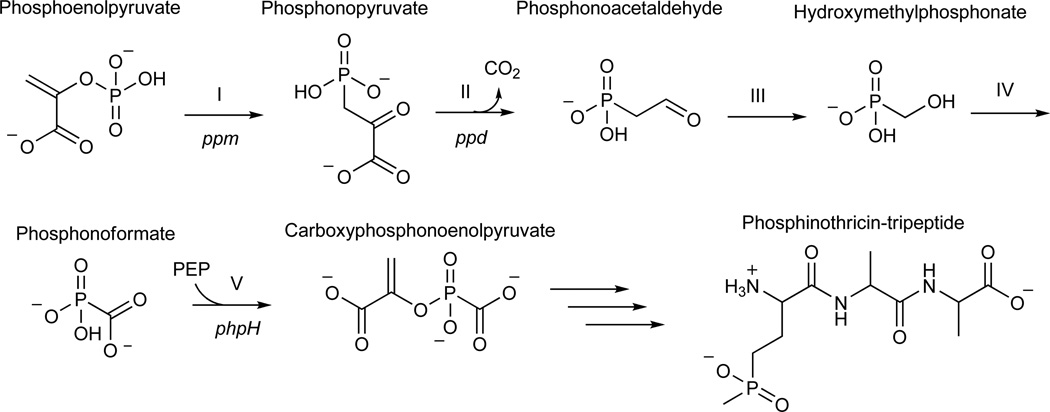
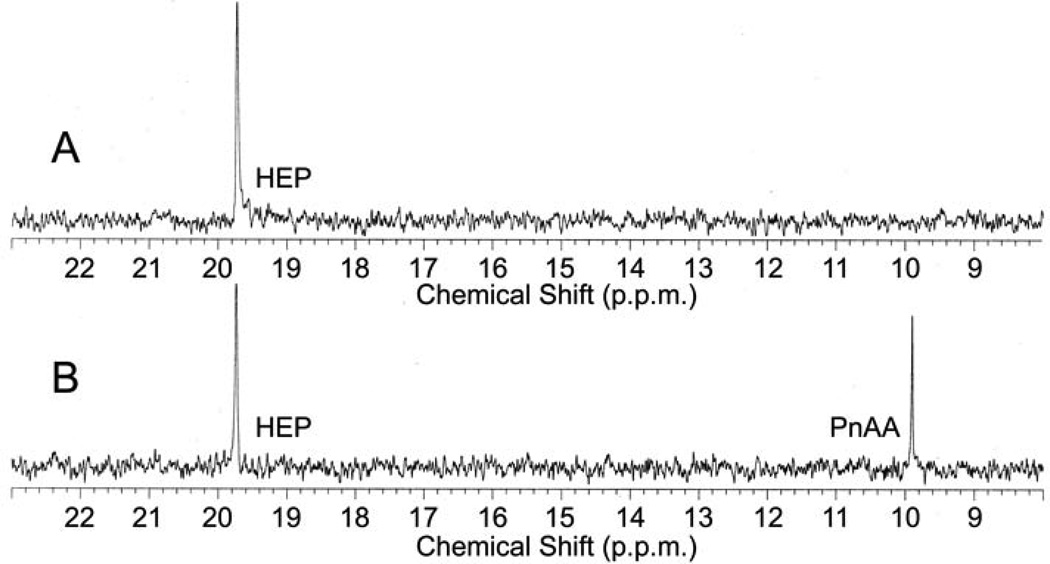
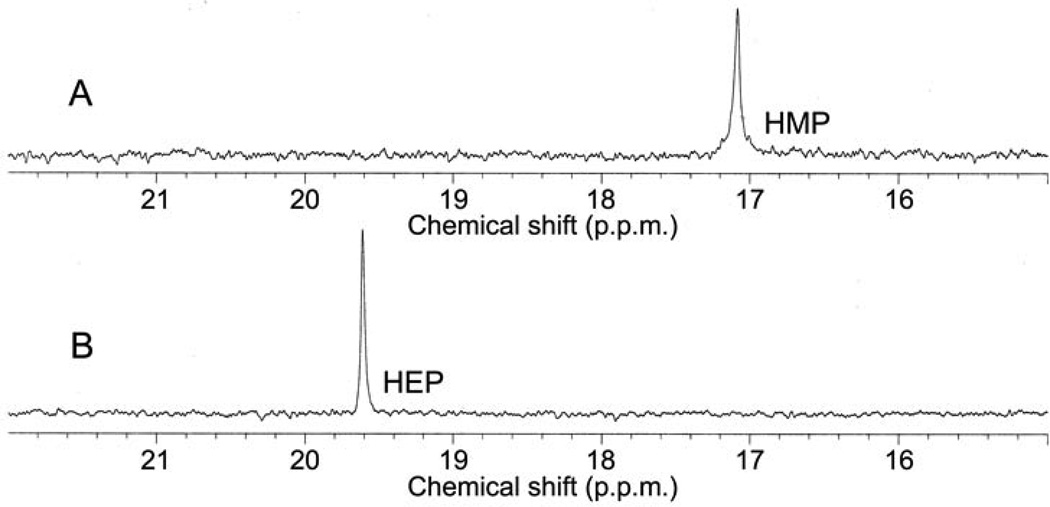
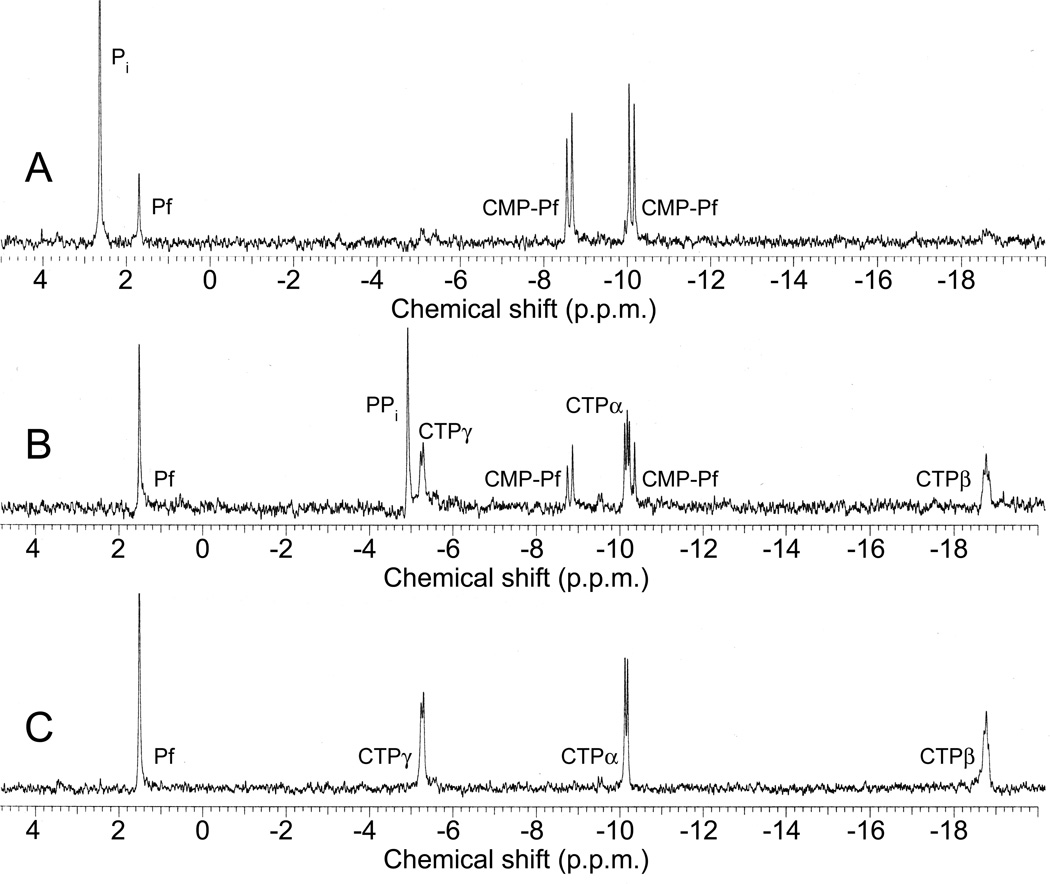
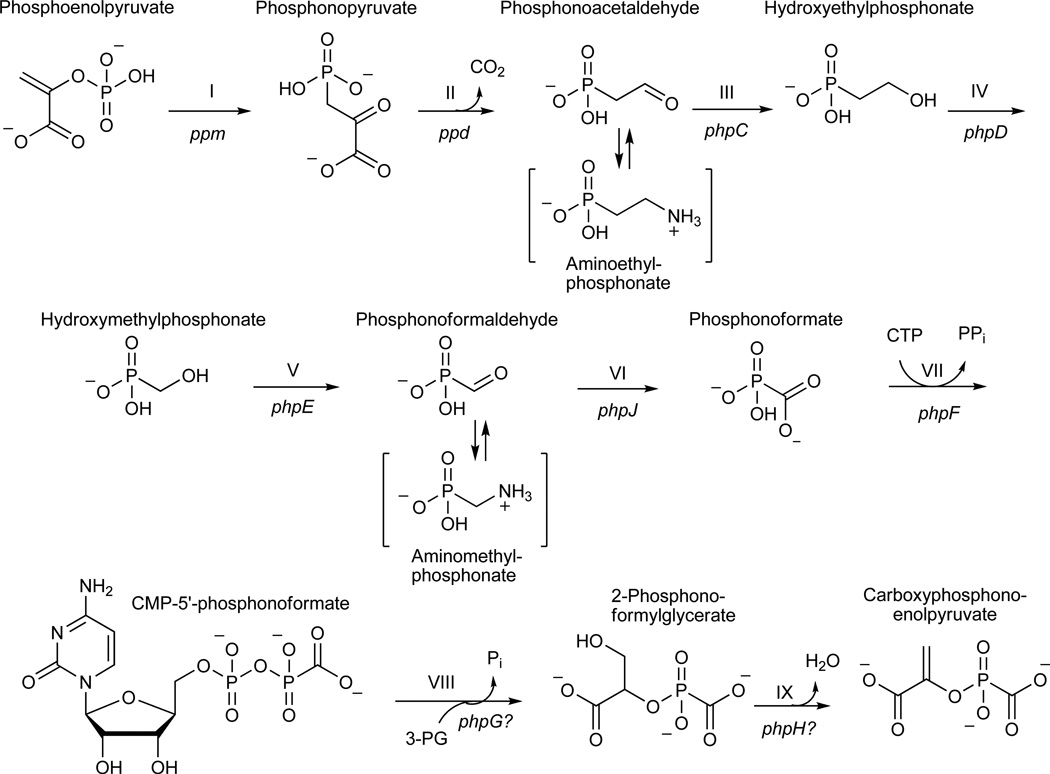
Phosphinothricin tripeptide (PTT, phosphinothricylalanylalanine) is a natural-product antibiotic and potent herbicide that is produced by Streptomyces hygroscopicus ATCC 21705 (ref. 1) and Streptomyces viridochromogenes DSM 40736 (ref. 2). PTT has attracted widespread interest because of its commercial applications and unique phosphinic acid functional group. Despite intensive study since its discovery in 1972 (see ref. 3 for a comprehensive review), a number of steps early in the PTT biosynthetic pathway remain uncharacterized. Here we report a series of interdisciplinary experiments involving the construction of defined S. viridochromogenes mutants, chemical characterization of accumulated intermediates, and in vitro assay of selected enzymes to examine these critical steps in PTT biosynthesis. Our results indicate that early PTT biosynthesis involves a series of catalytic steps that to our knowledge has not been described so far, including a highly unusual reaction for carbon bond cleavage. In sum, we define a pathway for early PTT biosynthesis that is more complex than previously appreciated.
Figures
References
-
- Seto H, et al. Studies on the biosynthesis of bialaphos (SF-1293). 1. Incorporation of 13C- and 2H-labeled precursors into bialaphos. The Journal of antibiotics. 1982;35(12):1719–1721. - PubMed
-
- Bayer E, et al. Phosphinothricin und Phosphinothricyl-Alanyl-Alanin. Helvetica Chimica Acta. 1972;55:224–239. - PubMed
-
- Thompson CJ, Seto H. In: Genetics and Biochemistry of Antibiotic Production. Vining LC, Stuttard C, editors. Newton, MA: Butterworth- Heinemann; 1995. pp. 197–222.
-
- Abell LM, Villafranca JJ. Investigation of the mechanism of phosphinothricin inactivation of Escherichia coli glutamine synthetase using rapid quench kinetic technique. Biochemistry. 1991;30:6135–6141. - PubMed
-
- Bailey RR. Brief overview of single-dose therapy for uncomplicated urinary tract infections. Chemotherapy. 1990;36(Suppl 1):27–30. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- PubChem-Substance/24775889
- PubChem-Substance/24775890
- PubChem-Substance/24775891
- PubChem-Substance/24775892
- PubChem-Substance/24775893
- PubChem-Substance/24775894
- PubChem-Substance/24775895
- PubChem-Substance/24775896
- PubChem-Substance/24775897
- PubChem-Substance/24775898
- PubChem-Substance/24775899
- PubChem-Substance/24775900
- PubChem-Substance/24775901
- PubChem-Substance/24775902
- PubChem-Substance/24775903
- PubChem-Substance/24775904
- PubChem-Substance/24775905
- PubChem-Substance/24775906
- PubChem-Substance/24775907
- PubChem-Substance/24775908
- PubChem-Substance/24775909
- PubChem-Substance/24775910
- PubChem-Substance/24775911
- PubChem-Substance/24775912
- PubChem-Substance/24775913
- PubChem-Substance/24775914
- PubChem-Substance/24775915
- PubChem-Substance/24775916
- PubChem-Substance/24775917
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
