

Host cell cathepsins potentiate Moloney murine leukemia virus infection
- PMID: 17634228
- PMCID: PMC2045468
- DOI: 10.1128/JVI.02853-06
Host cell cathepsins potentiate Moloney murine leukemia virus infection
Abstract
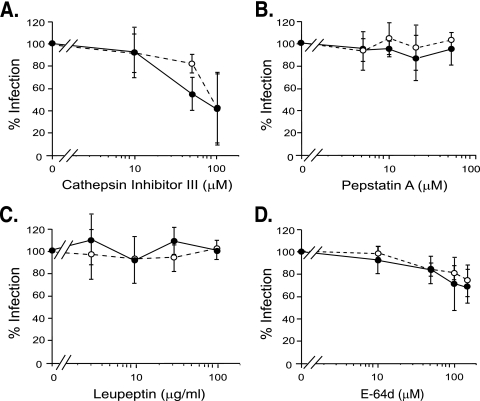
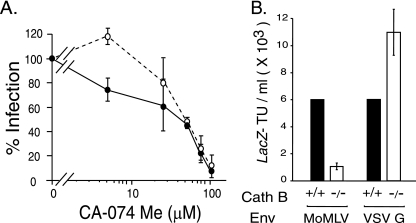
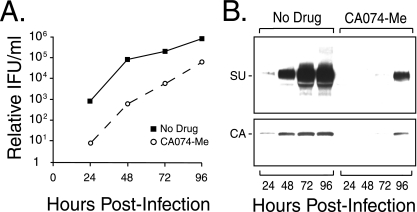
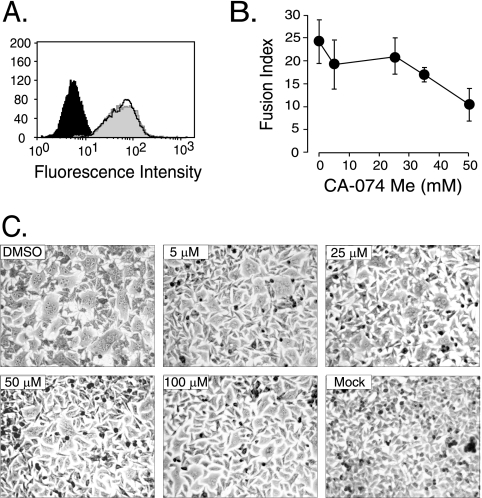
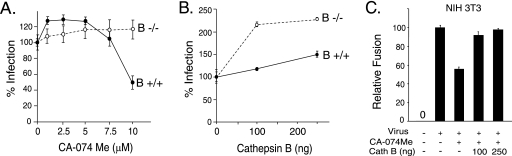
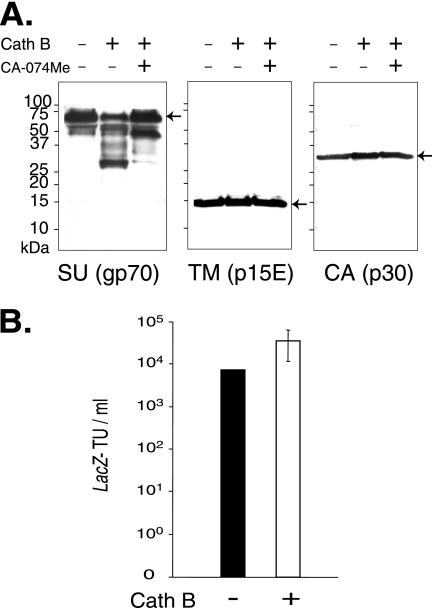
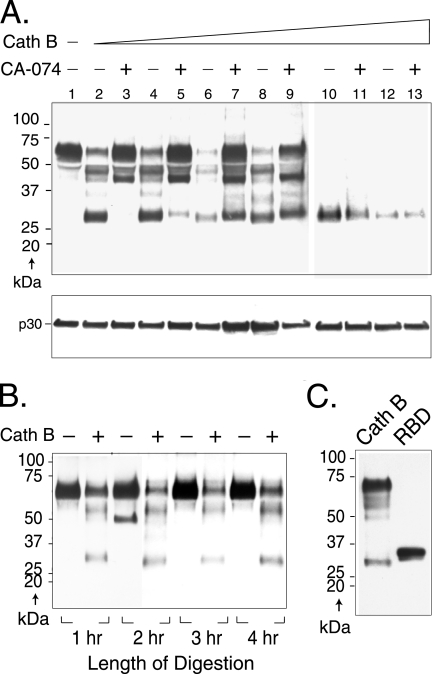
The roles of cellular proteases in Moloney murine leukemia virus (MLV) infection were investigated using MLV particles pseudotyped with vesicular stomatitis virus (VSV) G glycoprotein as a control for effects on core MLV particles versus effects specific to Moloney MLV envelope protein (Env). The broad-spectrum inhibitors cathepsin inhibitor III and E-64d gave comparable dose-dependent inhibition of Moloney MLV Env and VSV G pseudotypes, suggesting that the decrease did not involve the envelope protein. Whereas, CA-074 Me gave a biphasic response that differentiated between Moloney MLV Env and VSV G at low concentrations, at which the drug is highly selective for cathepsin B, but was similar for both glycoproteins at higher concentrations, at which CA-074 Me inhibits other cathepsins. Moloney MLV infection was lower on cathepsin B knockout fibroblasts than wild-type cells, whereas VSV G infection was not reduced on the B-/- cells. Taken together, these results support the notion that cathepsin B acts at an envelope-dependent step while another cathepsin acts at an envelope-independent step, such as uncoating or viral-DNA synthesis. Virus binding was not affected by CA-074 Me, whereas syncytium induction was inhibited in a dose-dependent manner, consistent with cathepsin B involvement in membrane fusion. Western blot analysis revealed specific cathepsin B cleavage of SU in vitro, while TM and CA remained intact. Infection could be enhanced by preincubation of Moloney MLV with cathepsin B, consistent with SU cleavage potentiating infection. These data suggested that during infection of NIH 3T3 cells, endocytosis brings Moloney MLV to early lysosomes, where the virus encounters cellular proteases, including cathepsin B, that cleave SU.
Figures
References
-
- Andersen, K. B. 1987. Cleavage fragments of the retrovirus surface protein gp70 during virus entry. J. Gen. Virol. 68:2193-2202. - PubMed
-
- Andersen, K. B., and B. A. Nexo. 1983. Entry of murine retrovirus into mouse fibroblasts. Virology 125:85-98. - PubMed
-
- Andersen, K. B., and H. Skov. 1989. Retrovirus-induced cell fusion is enhanced by protease treatment. J. Gen. Virol. 70:1921-1927. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
