

Inactivation of Rho GTPases with Clostridium difficile toxin B impairs centrosomal activation of Aurora-A in G2/M transition of HeLa cells
- PMID: 17634283
- PMCID: PMC1995717
- DOI: 10.1091/mbc.e07-03-0281
Inactivation of Rho GTPases with Clostridium difficile toxin B impairs centrosomal activation of Aurora-A in G2/M transition of HeLa cells
Abstract
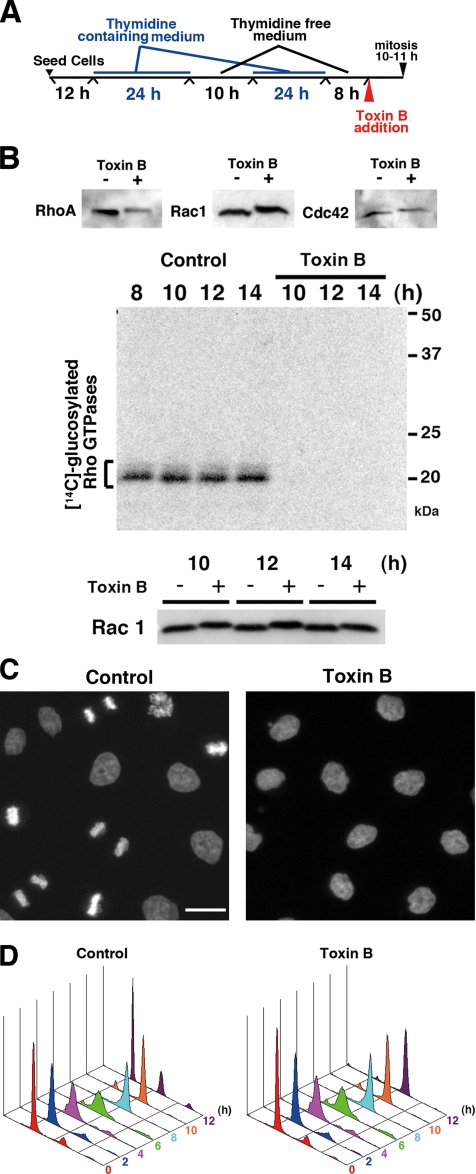
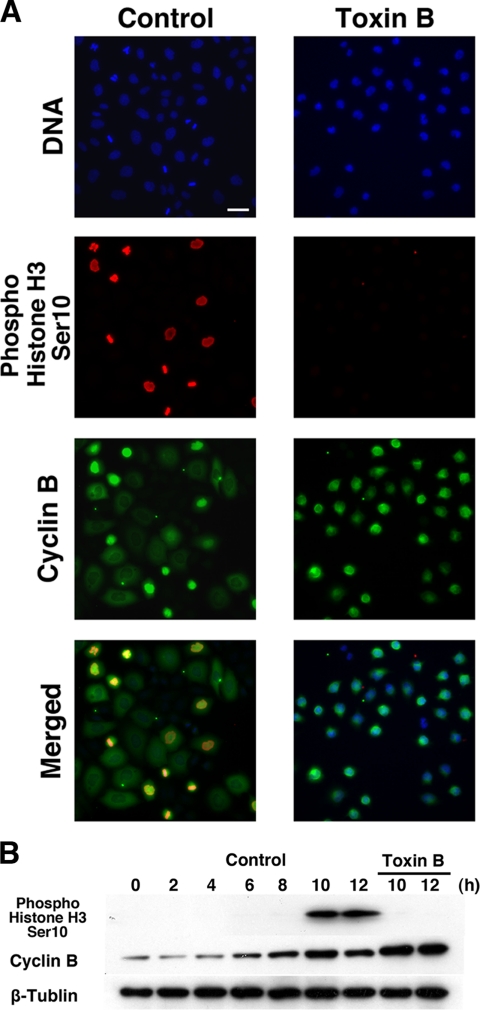
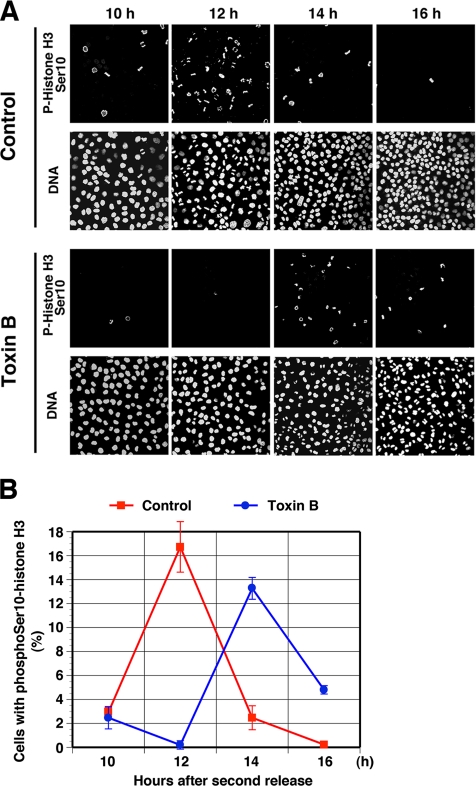
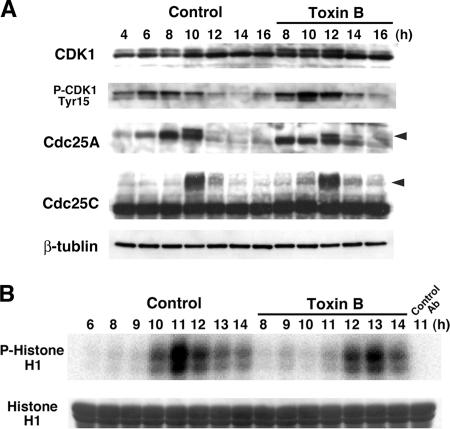
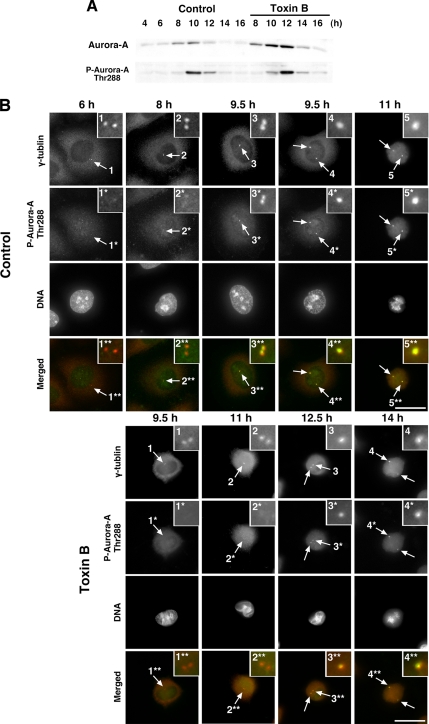
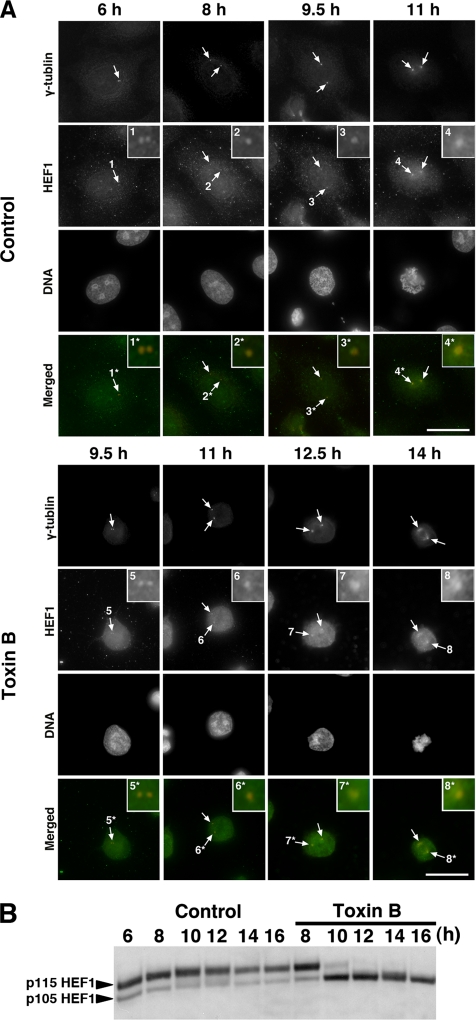
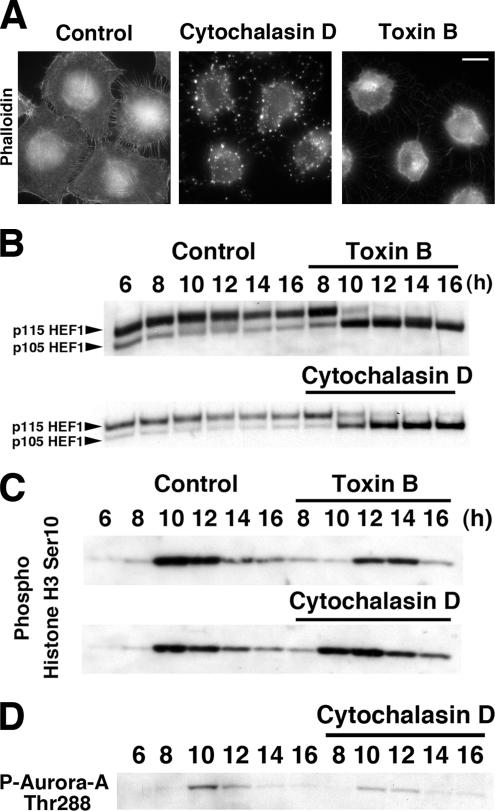
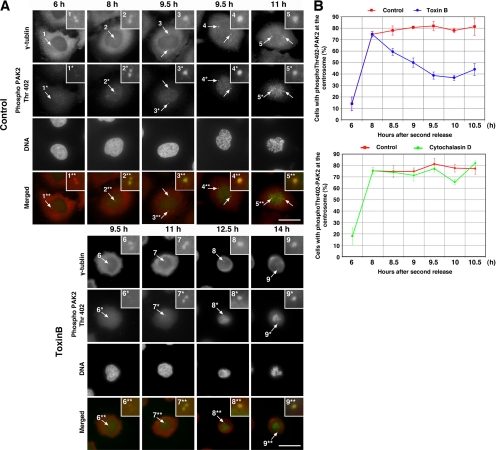
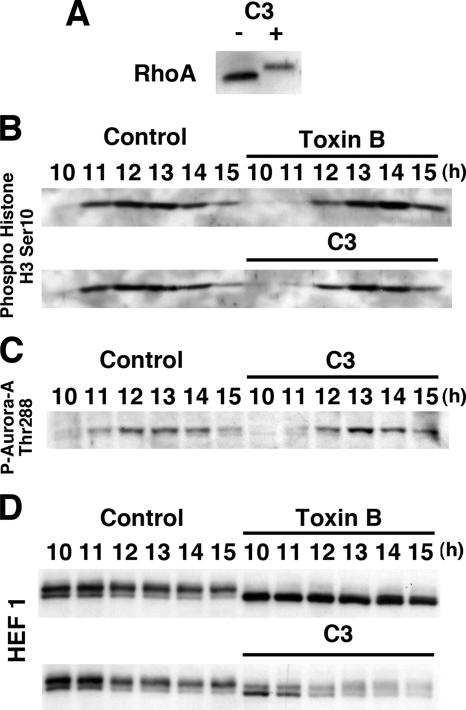
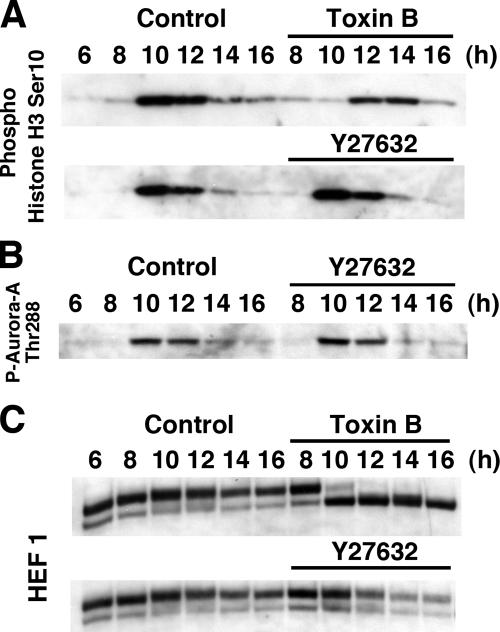
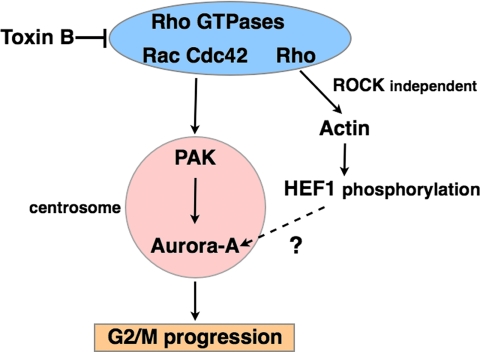
During G2 phase of cell cycle, centrosomes function as a scaffold for activation of mitotic kinases. Aurora-A is first activated at late G2 phase at the centrosome, facilitates centrosome maturation, and induces activation of cyclin B-Cdk1 at the centrosome for mitotic entry. Although several molecules including HEF1 and PAK are implicated in centrosomal activation of Aurora-A, signaling pathways leading to Aurora-A activation at the centrosome, and hence mitotic commitment in vertebrate cells remains largely unknown. Here, we have used Clostridium difficile toxin B and examined the role of Rho GTPases in G2/M transition of HeLa cells. Inactivation of Rho GTPases by the toxin B treatment delayed by 2 h histone H3 phosphorylation, Cdk1/cyclin B activation, and Aurora-A activation. Furthermore, PAK activation at the centrosome that was already present before the toxin addition was significantly attenuated for 2 h by the addition of toxin B, and HEF1 accumulation at the centrosome that occurred in late G2 phase was also delayed. These results suggest that Rho GTPases function in G2/M transition of mammalian cells by mediating multiple signaling pathways converging to centrosomal activation of Aurora-A.
Figures
References
-
- Aktories K., Barbieri J. T. Bacterial cytotoxins: targeting eukaryotic switches. Nat. Rev. Microbiol. 2005;3:397–410. - PubMed
-
- Bayliss R., Sardon T., Vernos I., Conti E. Structural basis of Aurora-A activation by TPX2 at the mitotic spindle. Mol. Cell. 2003;12:851–862. - PubMed
-
- Belmont A. S. Mitotic chromosome structure and condensation. Curr. Opin. Cell Biol. 2006;18:632–638. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
