

D1 dopamine receptor dDA1 is required in the mushroom body neurons for aversive and appetitive learning in Drosophila
- PMID: 17634358
- PMCID: PMC6672866
- DOI: 10.1523/JNEUROSCI.1167-07.2007
D1 dopamine receptor dDA1 is required in the mushroom body neurons for aversive and appetitive learning in Drosophila
Abstract
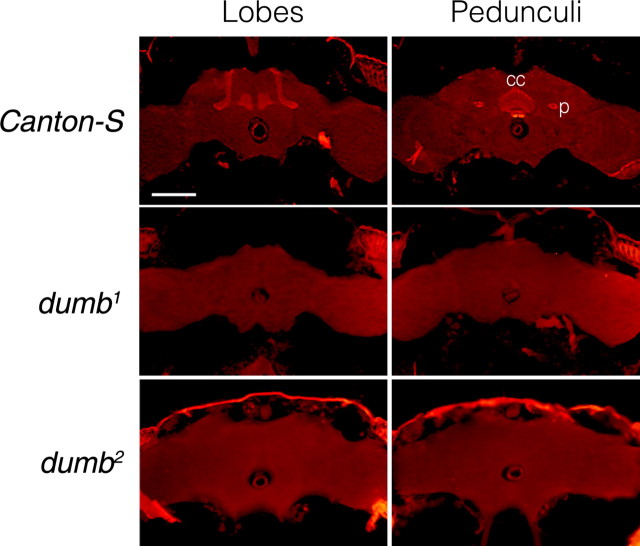
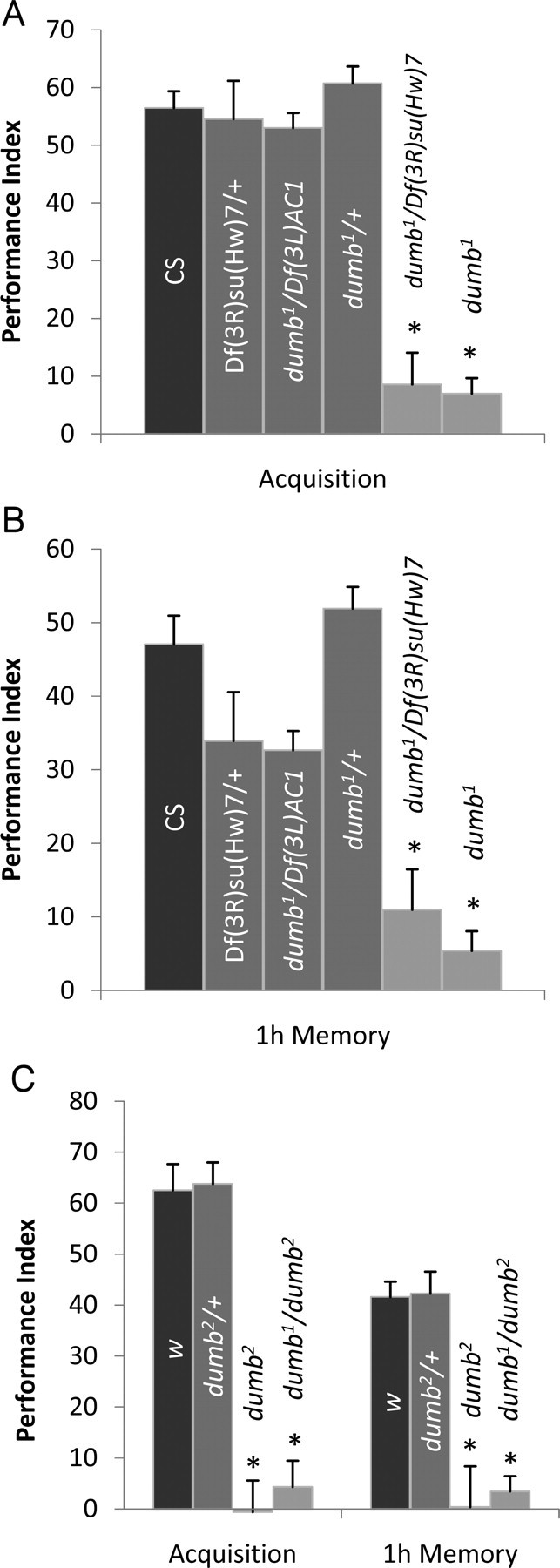
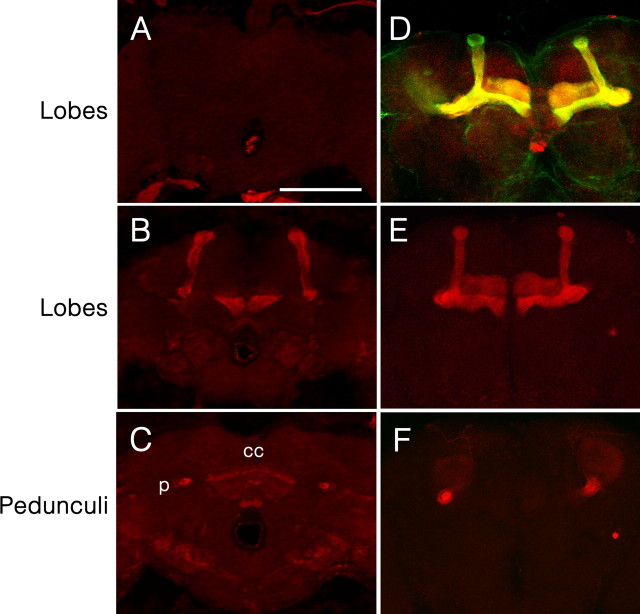
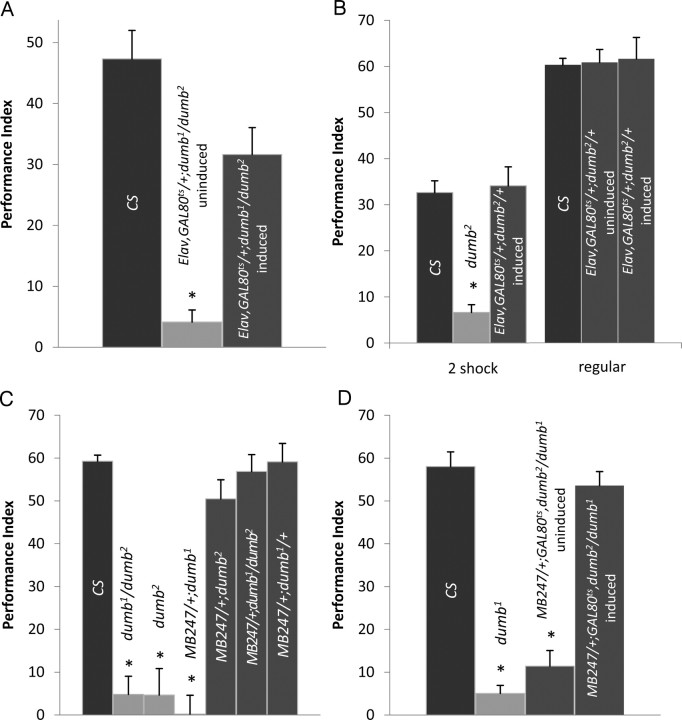
Drosophila has robust behavioral plasticity to avoid or prefer the odor that predicts punishment or food reward, respectively. Both types of plasticity are mediated by the mushroom body (MB) neurons in the brain, in which various signaling molecules play crucial roles. However, important yet unresolved molecules are the receptors that initiate aversive or appetitive learning cascades in the MB. We have shown previously that D1 dopamine receptor dDA1 is highly enriched in the MB neuropil. Here, we demonstrate that dDA1 is a key receptor that mediates both aversive and appetitive learning in pavlovian olfactory conditioning. We identified two mutants, dumb1 and dumb2, with abnormal dDA1 expression. When trained with the same conditioned stimuli, both dumb alleles showed negligible learning in electric shock-mediated conditioning while they exhibited moderately impaired learning in sugar-mediated conditioning. These phenotypes were not attributable to anomalous sensory modalities of dumb mutants because their olfactory acuity, shock reactivity, and sugar preference were comparable to those of control lines. Remarkably, the dumb mutant's impaired performance in both paradigms was fully rescued by reinstating dDA1 expression in the same subset of MB neurons, indicating the critical roles of the MB dDA1 in aversive as well as appetitive learning. Previous studies using dopamine receptor antagonists implicate the involvement of D1/D5 receptors in various pavlovian conditioning tasks in mammals; however, these have not been supported by the studies of D1- or D5-deficient animals. The findings described here unambiguously clarify the critical roles of D1 dopamine receptor in aversive and appetitive pavlovian conditioning.
Figures
Similar articles
-
Concerted Actions of Octopamine and Dopamine Receptors Drive Olfactory Learning.J Neurosci. 2020 May 20;40(21):4240-4250. doi: 10.1523/JNEUROSCI.1756-19.2020. Epub 2020 Apr 10. J Neurosci. 2020. PMID: 32277043 Free PMC article.
-
Dopamine is required for learning and forgetting in Drosophila.Neuron. 2012 May 10;74(3):530-42. doi: 10.1016/j.neuron.2012.04.007. Neuron. 2012. PMID: 22578504 Free PMC article.
-
Distinctive neuronal networks and biochemical pathways for appetitive and aversive memory in Drosophila larvae.J Neurosci. 2009 Jan 21;29(3):852-62. doi: 10.1523/JNEUROSCI.1315-08.2009. J Neurosci. 2009. PMID: 19158309 Free PMC article.
-
Olfactory learning in Drosophila.Physiology (Bethesda). 2010 Dec;25(6):338-46. doi: 10.1152/physiol.00026.2010. Physiology (Bethesda). 2010. PMID: 21186278 Free PMC article. Review.
-
Roles of Octopamine and Dopamine Neurons for Mediating Appetitive and Aversive Signals in Pavlovian Conditioning in Crickets.Front Physiol. 2017 Dec 12;8:1027. doi: 10.3389/fphys.2017.01027. eCollection 2017. Front Physiol. 2017. PMID: 29311961 Free PMC article. Review.
Cited by
-
Identification of a dopamine pathway that regulates sleep and arousal in Drosophila.Nat Neurosci. 2012 Nov;15(11):1516-23. doi: 10.1038/nn.3238. Epub 2012 Oct 14. Nat Neurosci. 2012. PMID: 23064381
-
Associative learning drives longitudinally graded presynaptic plasticity of neurotransmitter release along axonal compartments.Elife. 2022 Mar 14;11:e76712. doi: 10.7554/eLife.76712. Elife. 2022. PMID: 35285796 Free PMC article.
-
Dopamine activity in projection neurons regulates short-lasting olfactory approach memory in Drosophila.Eur J Neurosci. 2022 Sep;56(5):4558-4571. doi: 10.1111/ejn.15766. Epub 2022 Jul 17. Eur J Neurosci. 2022. PMID: 35815601 Free PMC article.
-
Differential second messenger signaling via dopamine neurons bidirectionally regulates memory retention.Proc Natl Acad Sci U S A. 2023 Sep 5;120(36):e2304851120. doi: 10.1073/pnas.2304851120. Epub 2023 Aug 28. Proc Natl Acad Sci U S A. 2023. PMID: 37639608 Free PMC article.
-
Functional architecture of reward learning in mushroom body extrinsic neurons of larval Drosophila.Nat Commun. 2018 Mar 16;9(1):1104. doi: 10.1038/s41467-018-03130-1. Nat Commun. 2018. PMID: 29549237 Free PMC article.
References
-
- Baker RM, Shah MJ, Sclafani A, Bodnar RJ. Dopamine D1 and D2 antagonists reduce the acquisition and expression of flavor-preferences conditioned by fructose in rats. Pharmacol Biochem Behav. 2003;75:55–65. - PubMed
-
- Budnik V, White K. Genetic dissection of dopamine and serotonin synthesis in the nervous system of Drosophila melanogaster. J Neurogenet. 1987;4:309–314. - PubMed
-
- Cannon CM, Scannell CA, Palmiter RD. Mice lacking dopamine D1 receptors express normal lithium chloride-induced conditioned taste aversion for salt but not sucrose. Eur J Neurosci. 2005;21:2600–2604. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous