

Presynaptic contributions of chordin to hippocampal plasticity and spatial learning
- PMID: 17634368
- PMCID: PMC6672865
- DOI: 10.1523/JNEUROSCI.1604-07.2007
Presynaptic contributions of chordin to hippocampal plasticity and spatial learning
Abstract
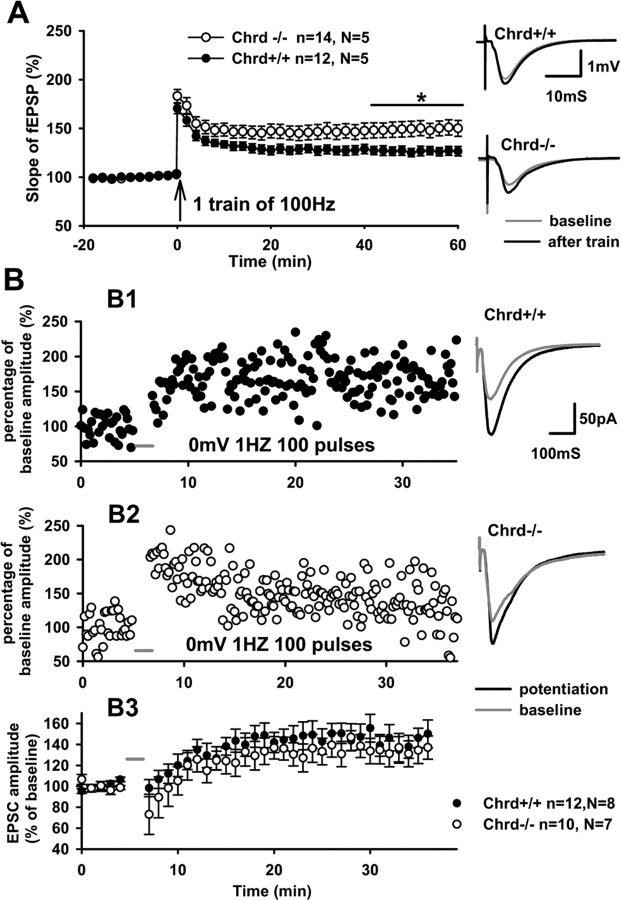
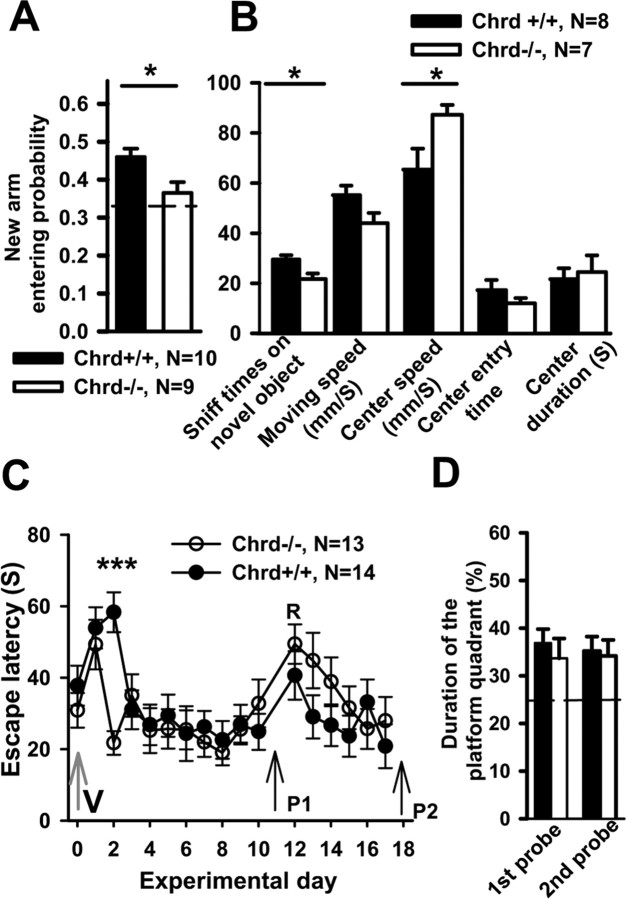
Recently, several evolutionary conserved signaling pathways that play prominent roles in regulating early neurodevelopment have been found to regulate synaptic remodeling in the adult. To test whether adult neuronal expression of bone morphogenic protein (BMP) signaling components also plays a postnatal role in regulating neuronal plasticity, we modulated BMP signaling in mice both in vivo and in vitro by genetic removal of the BMP inhibitor chordin or by perfusing recombinant BMP signaling pathway components onto acute hippocampal slices. Chordin null mice exhibited a significant increase in presynaptic transmitter release from hippocampal neurons, resulting in enhanced paired-pulse facilitation and long-term potentiation. These mice also showed a decreased acquisition time in a water maze test along with less exploratory activity during Y-maze and open-field tests. Perfusion of BMP ligands onto hippocampal slices replicated the presynaptic phenotype of chordin null slices, but bath application of Noggin, another antagonist of BMP signaling pathway, significantly decrease the frequency of miniature EPSCs. These results demonstrate that the BMP signaling pathway contributes to synaptic plasticity and learning likely through a presynaptic mechanism.
Figures






References
-
- Abbott LF, Regehr WG. Synaptic computation. Nature. 2004;431:796–803. - PubMed
-
- Aberle H, Haghighi AP, Fetter RD, McCabe BD, Magalhaes TR, Goodman CS. wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila . Neuron. 2002;33:545–558. - PubMed
-
- Bachiller D, Klingensmith J, Kemp C, Belo JA, Anderson RM, May SR, McMahon JA, McMahon AP, Harland RM, Rossant J, De Robertis EM. The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature. 2000;403:658–661. - PubMed
-
- Bachiller D, Klingensmith J, Shneyder N, Tran U, Anderson R, Rossant J, De Robertis EM. The role of chordin/Bmp signals in mammalian pharyngeal development and DiGeorge syndrome. Development. 2003;130:3567–3578. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases