

Physiologically distinct temporal cohorts of cortical interneurons arise from telencephalic Olig2-expressing precursors
- PMID: 17634372
- PMCID: PMC6672881
- DOI: 10.1523/JNEUROSCI.1807-07.2007
Physiologically distinct temporal cohorts of cortical interneurons arise from telencephalic Olig2-expressing precursors
Abstract
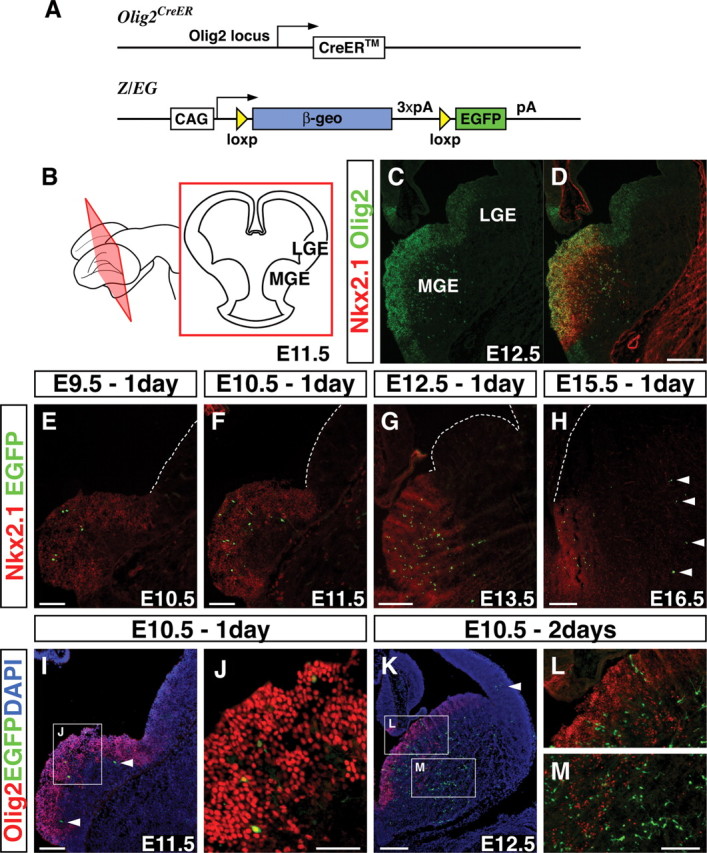
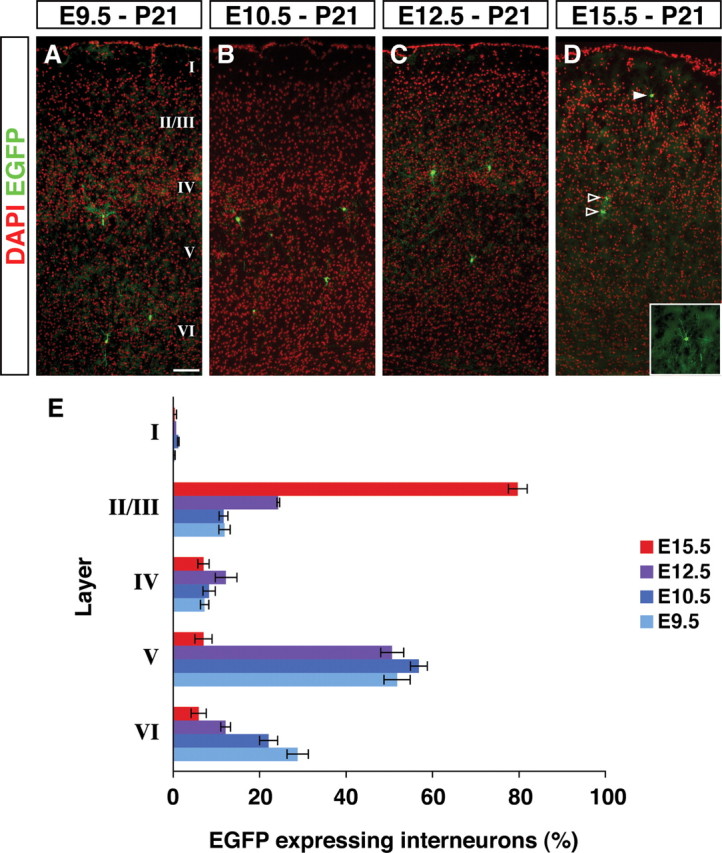
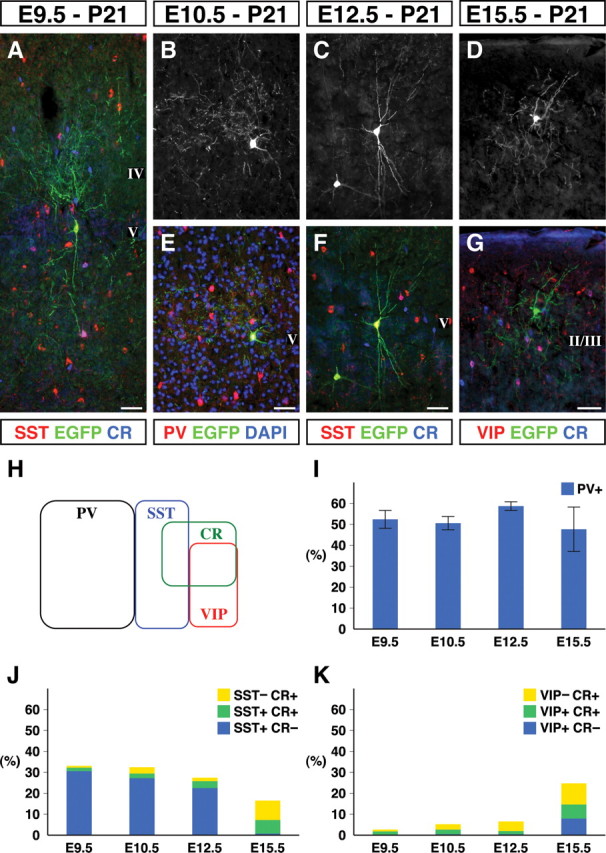
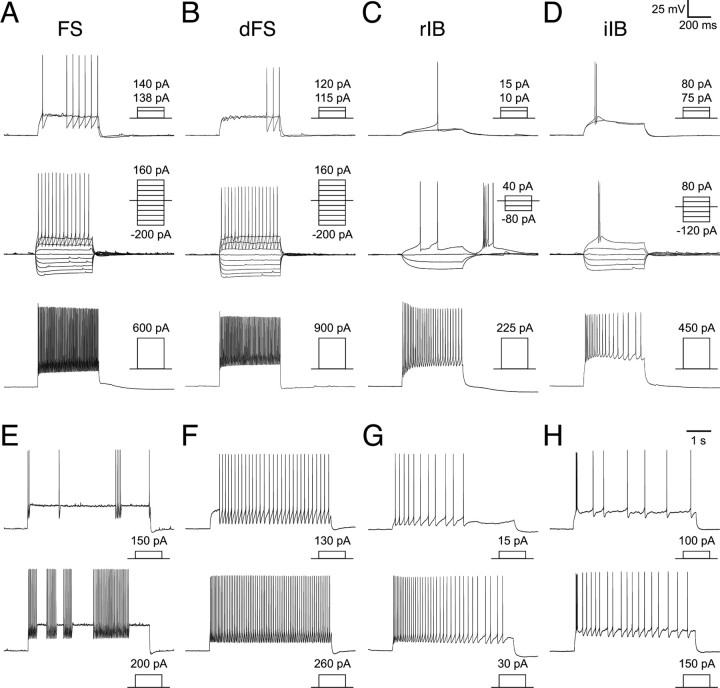
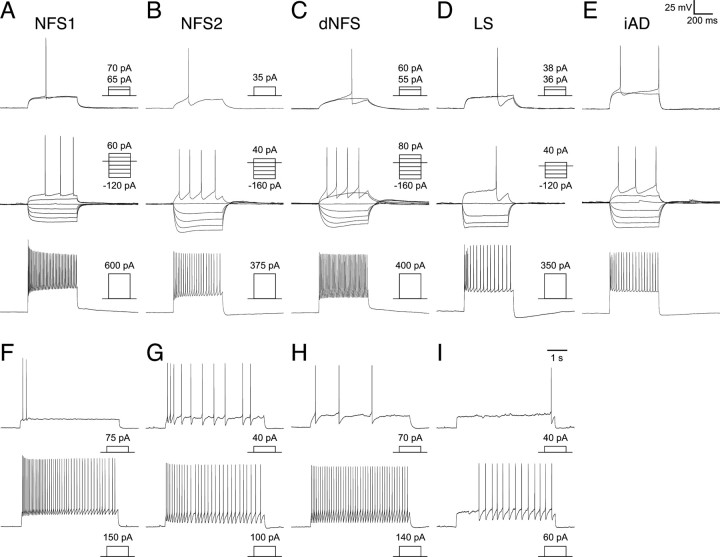
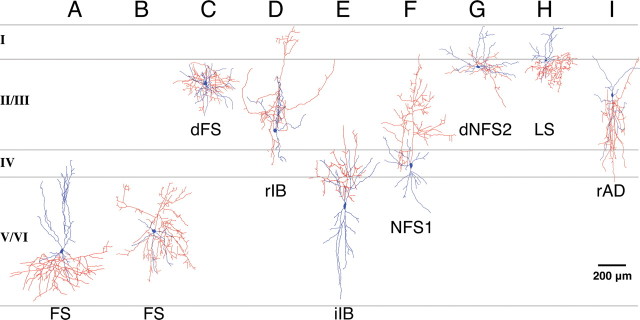
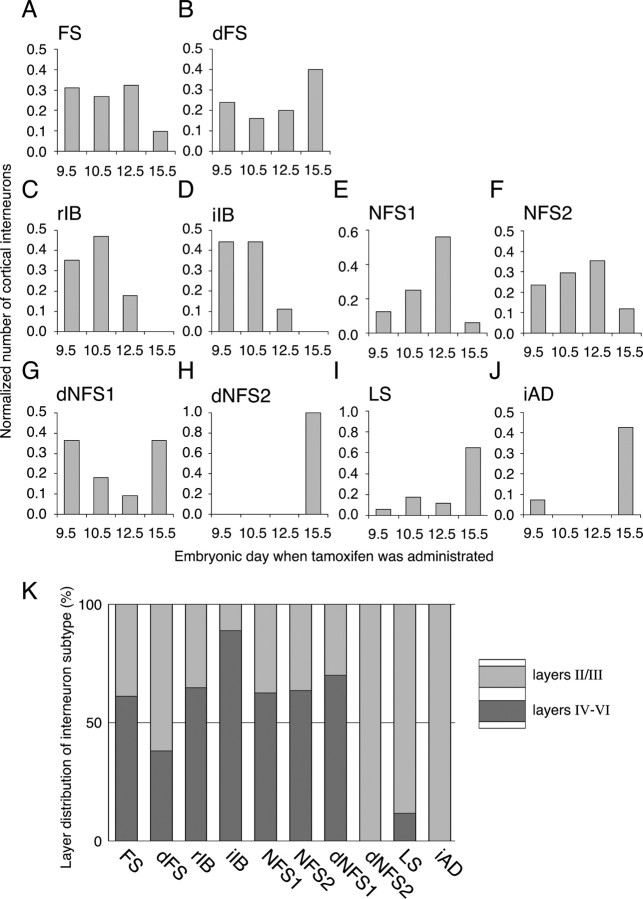
Inhibitory GABAergic interneurons of the mouse neocortex are a highly heterogeneous population of neurons that originate from the ventral telencephalon and migrate tangentially up into the developing cortical plate. The majority of cortical interneurons arise from a transient embryonic structure known as the medial ganglionic eminence (MGE), but how the remarkable diversity is specified in this region is not known. We have taken a genetic fate mapping strategy to elucidate the temporal origins of cortical interneuron subtypes within the MGE. We used an inducible form of Cre under the regulation of Olig2, a basic helix-loop-helix transcription factor highly expressed in neural progenitors of the MGE. We observe that the physiological subtypes of cortical interneurons are, to a large degree, unique to their time point of generation.
Figures
Similar articles
-
RhoA and Cdc42 are required in pre-migratory progenitors of the medial ganglionic eminence ventricular zone for proper cortical interneuron migration.Development. 2013 Aug;140(15):3139-45. doi: 10.1242/dev.092585. Development. 2013. PMID: 23861058 Free PMC article.
-
A blueprint for the spatiotemporal origins of mouse hippocampal interneuron diversity.J Neurosci. 2011 Jul 27;31(30):10948-70. doi: 10.1523/JNEUROSCI.0323-11.2011. J Neurosci. 2011. PMID: 21795545 Free PMC article.
-
The requirement of Nkx2-1 in the temporal specification of cortical interneuron subtypes.Neuron. 2008 Sep 11;59(5):722-32. doi: 10.1016/j.neuron.2008.07.031. Neuron. 2008. PMID: 18786356 Free PMC article.
-
Perspectives on the developmental origins of cortical interneuron diversity.Novartis Found Symp. 2007;288:21-35; discussion 35-44, 96-8. Novartis Found Symp. 2007. PMID: 18494250 Review.
-
The origin and specification of cortical interneurons.Nat Rev Neurosci. 2006 Sep;7(9):687-96. doi: 10.1038/nrn1954. Epub 2006 Aug 2. Nat Rev Neurosci. 2006. PMID: 16883309 Review.
Cited by
-
Human embryonic stem cell-derived oligodendrocytes: protocols and perspectives.Stem Cells Dev. 2013 Sep 15;22(18):2459-76. doi: 10.1089/scd.2012.0520. Epub 2013 Jun 12. Stem Cells Dev. 2013. PMID: 23621561 Free PMC article. Review.
-
Cell-type specific transcriptomic signatures of neocortical circuit organization and their relevance to autism.Front Neural Circuits. 2022 Sep 23;16:982721. doi: 10.3389/fncir.2022.982721. eCollection 2022. Front Neural Circuits. 2022. PMID: 36213201 Free PMC article.
-
N-cadherin (Cdh2) Maintains Migration and Postmitotic Survival of Cortical Interneuron Precursors in a Cell-Type-Specific Manner.Cereb Cortex. 2020 Mar 14;30(3):1318-1329. doi: 10.1093/cercor/bhz168. Cereb Cortex. 2020. PMID: 31402374 Free PMC article.
-
Arx Expression Suppresses Ventralization of the Developing Dorsal Forebrain.Sci Rep. 2019 Jan 18;9(1):226. doi: 10.1038/s41598-018-36194-6. Sci Rep. 2019. PMID: 30659230 Free PMC article.
-
Many specialists for suppressing cortical excitation.Front Neurosci. 2008 Dec 15;2(2):155-67. doi: 10.3389/neuro.01.026.2008. eCollection 2008 Dec. Front Neurosci. 2008. PMID: 19225588 Free PMC article.
References
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Angevine JB, II, Sidman RL. Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature. 1961;192:766–768. - PubMed
-
- Bertrand N, Castro DS, Guillemot F. Proneural genes and the specification of neural cell types. Nat Rev Neurosci. 2002;3:517–530. - PubMed
-
- Butt SJ, Fuccillo M, Nery S, Noctor S, Kriegstein A, Corbin JG, Fishell G. The temporal and spatial origins of cortical interneurons predict their physiological subtype. Neuron. 2005;48:591–604. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases