

The secreted beta-amyloid precursor protein ectodomain APPs alpha is sufficient to rescue the anatomical, behavioral, and electrophysiological abnormalities of APP-deficient mice
- PMID: 17634375
- PMCID: PMC6672885
- DOI: 10.1523/JNEUROSCI.1026-07.2007
The secreted beta-amyloid precursor protein ectodomain APPs alpha is sufficient to rescue the anatomical, behavioral, and electrophysiological abnormalities of APP-deficient mice
Abstract
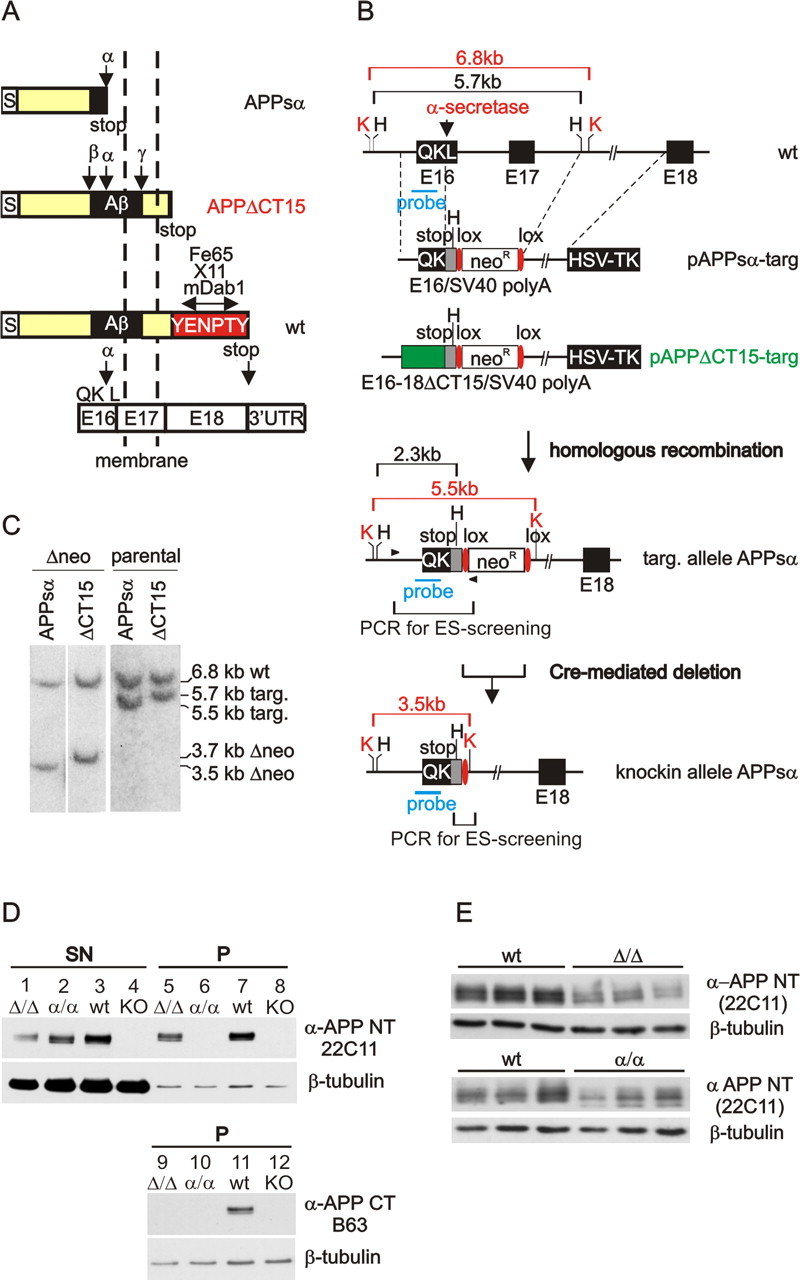
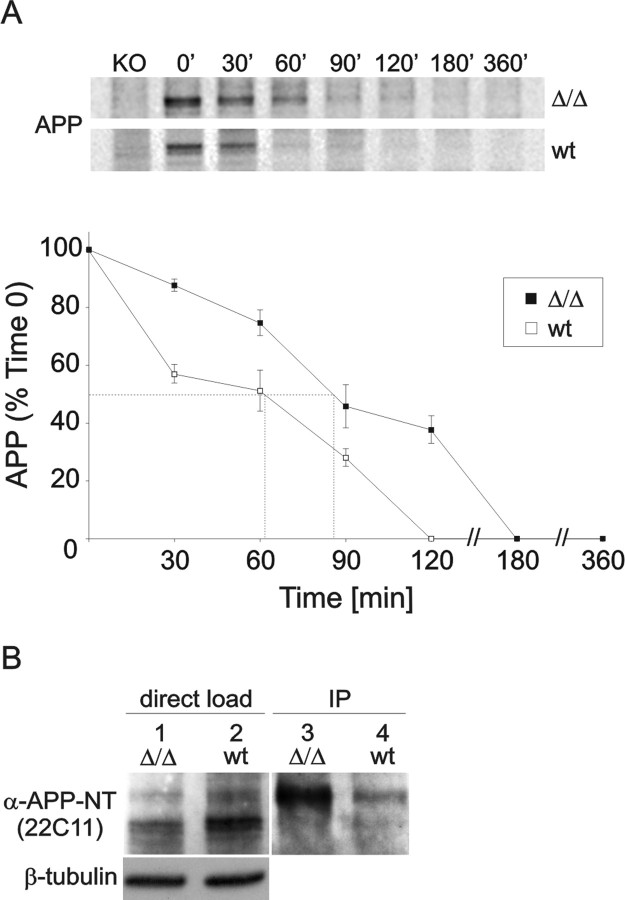
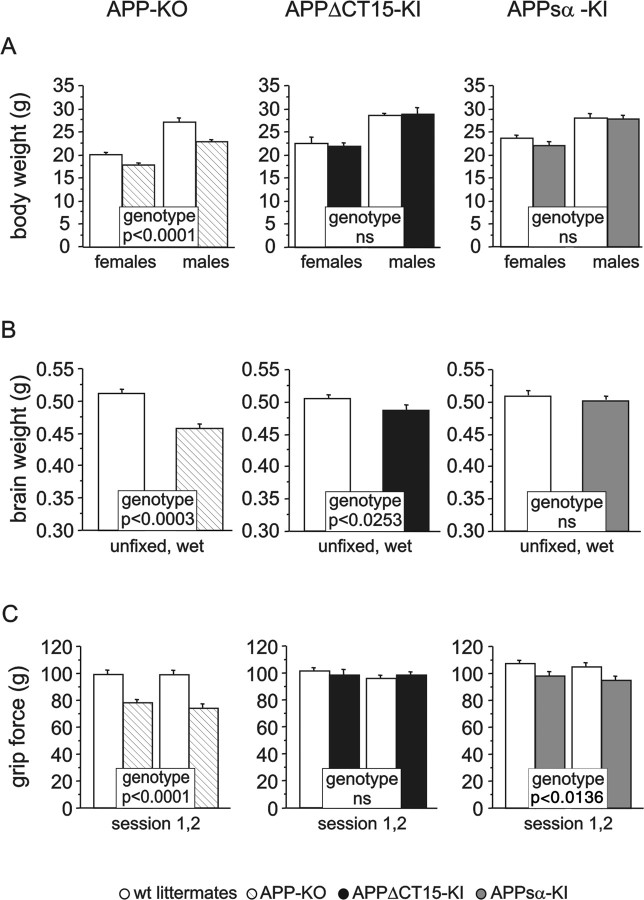
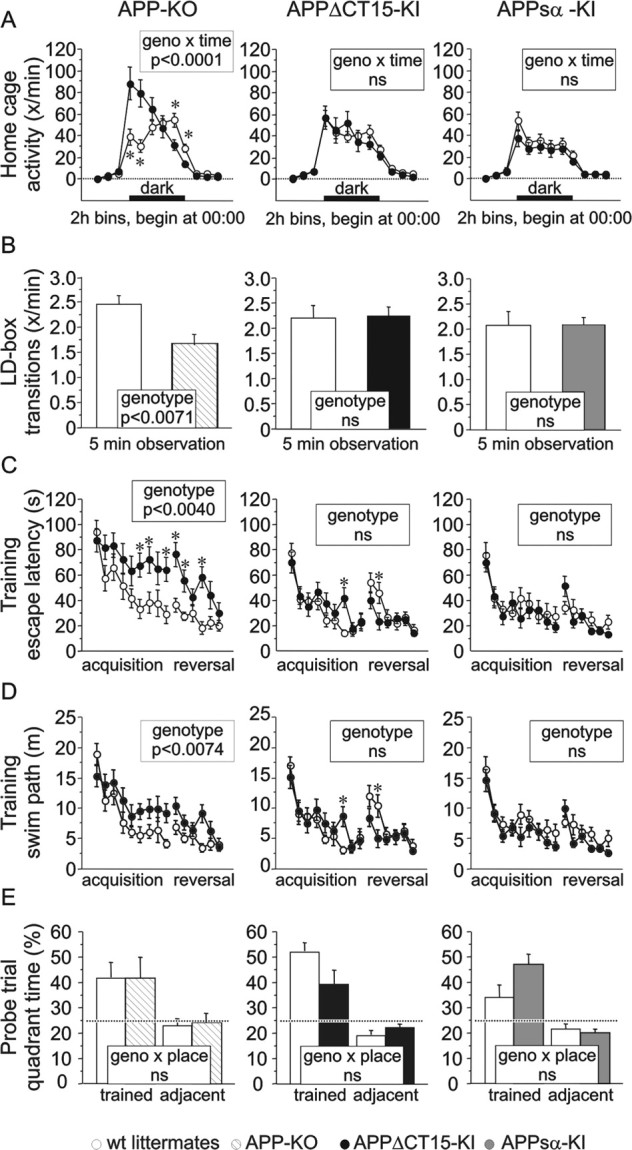
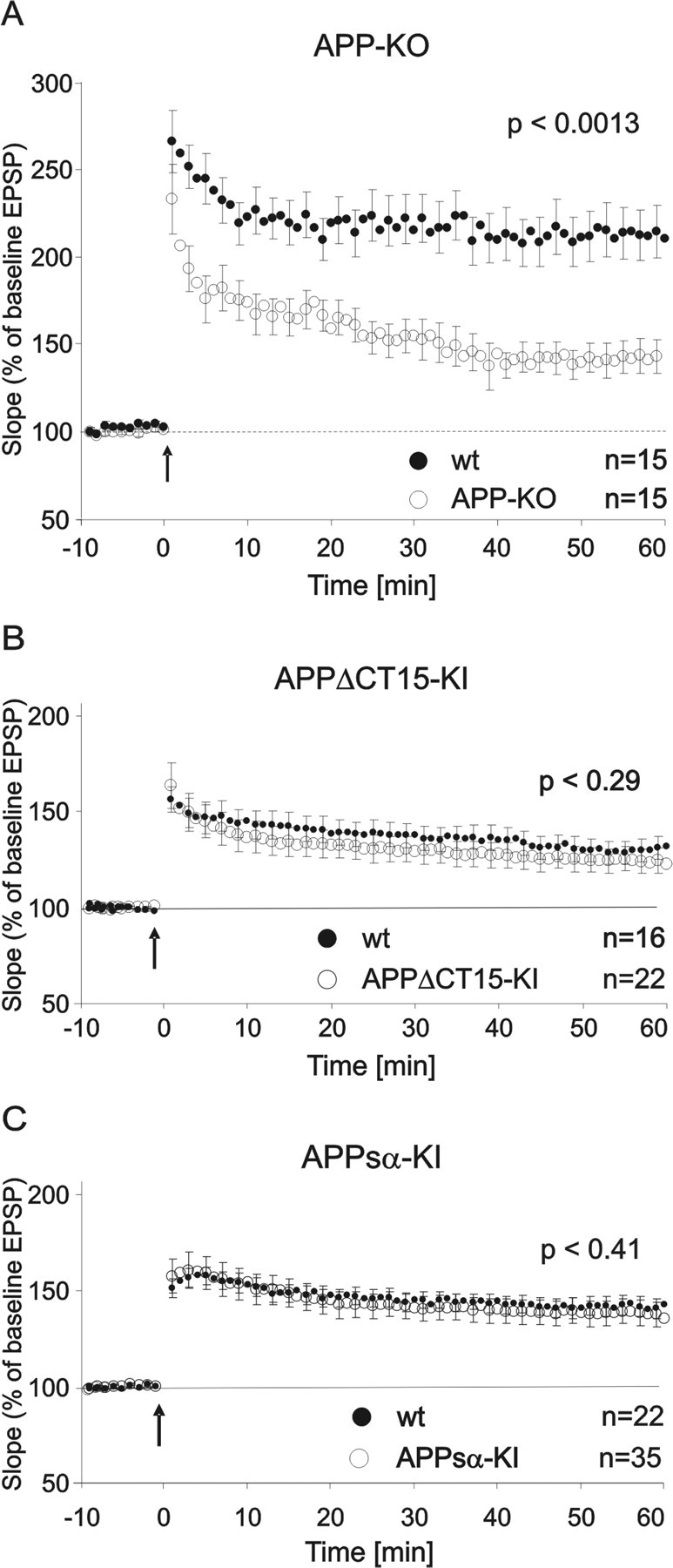
It is well established that the proteolytic processing of the beta-amyloid precursor protein (APP) generates beta-amyloid (Abeta), which plays a central role in the pathogenesis of Alzheimer's disease (AD). In contrast, the physiological role of APP and of its numerous proteolytic fragments and the question of whether a loss of these functions contributes to AD are still unknown. To address this question, we replaced the endogenous APP locus by gene-targeted alleles and generated two lines of knock-in mice that exclusively express APP deletion variants corresponding either to the secreted APP ectodomain (APPs alpha) or to a C-terminal (CT) truncation lacking the YENPTY interaction motif (APPdeltaCT15). Interestingly, the deltaCT15 deletion resulted in reduced turnover of holoAPP, increased cell surface expression, and strongly reduced Abeta levels in brain, likely because of reduced processing in the endocytic pathway. Most importantly, we demonstrate that in both APP knock-in lines the expression of APP N-terminal domains either grossly attenuated or completely rescued the prominent deficits of APP knock-out mice, such as reductions in brain and body weight, grip strength deficits, alterations in circadian locomotor activity, exploratory activity, and the impairment in spatial learning and long-term potentiation. Together, our data suggest that the APP C terminus is dispensable and that APPs alpha is sufficient to mediate the physiological functions of APP assessed by these tests.
Figures
References
-
- Almkvist O, Basun H, Wagner SL, Rowe BA, Wahlund LO, Lannfelt L. Cerebrospinal fluid levels of alpha-secretase-cleaved soluble amyloid precursor protein mirror cognition in a Swedish family with Alzheimer disease and a gene mutation. Arch Neurol. 1997;54:641–644. - PubMed
-
- Anderson JJ, Holtz G, Baskin PP, Wang R, Mazzarelli L, Wagner SL, Menzaghi F. Reduced cerebrospinal fluid levels of alpha-secretase-cleaved amyloid precursor protein in aged rats: correlation with spatial memory deficits. Neuroscience. 1999;93:1409–1420. - PubMed
-
- Anliker B, Müller U. The functions of the mammalian amyloid precursor protein and related amyloid precursor-like proteins. Neurodegener Dis. 2006;3:239–246. - PubMed
-
- Bell KF, Zheng L, Fahrenholz F, Cuello AC. ADAM-10 over-expression increases cortical synaptogenesis. Neurobiol Aging. 2007 in press. - PubMed
-
- Bour A, Little S, Dodart JC, Kelche C, Mathis C. A secreted form of the beta-amyloid precursor protein (sAPP695) improves spatial recognition memory in OF1 mice. Neurobiol Learn Mem. 2004;81:27–38. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases