

Pharmacological imposition of sleep slows cognitive decline and reverses dysregulation of circadian gene expression in a transgenic mouse model of Huntington's disease
- PMID: 17634381
- PMCID: PMC6672877
- DOI: 10.1523/JNEUROSCI.0649-07.2007
Pharmacological imposition of sleep slows cognitive decline and reverses dysregulation of circadian gene expression in a transgenic mouse model of Huntington's disease
Abstract
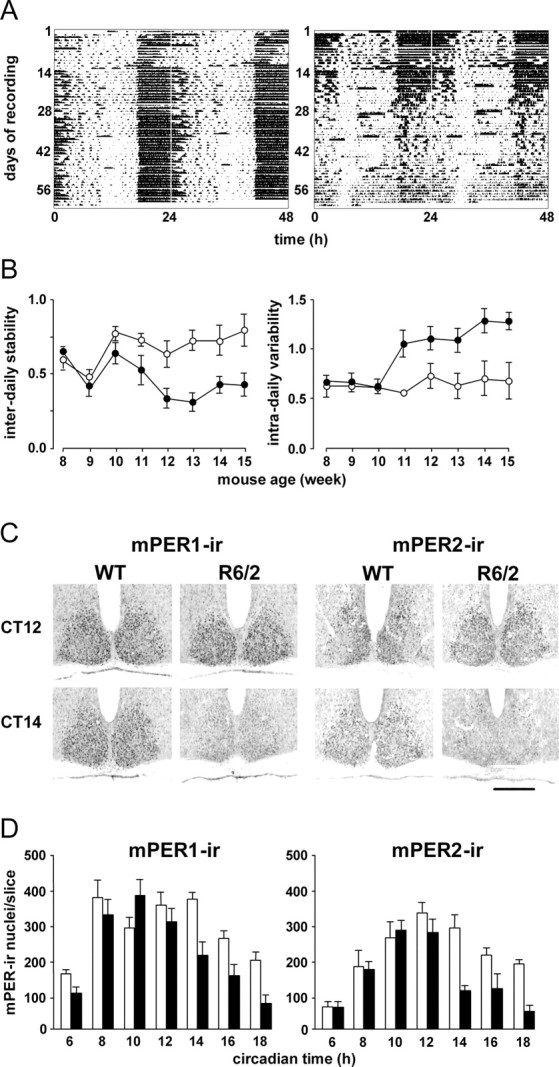
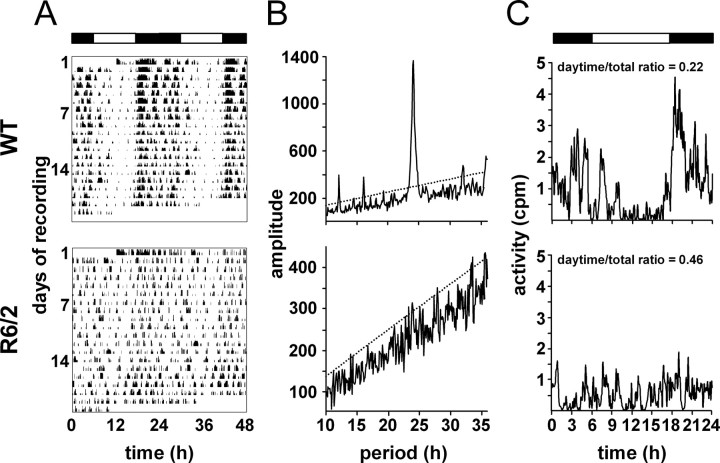
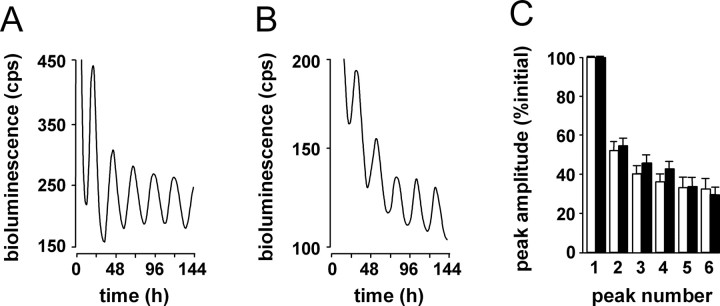
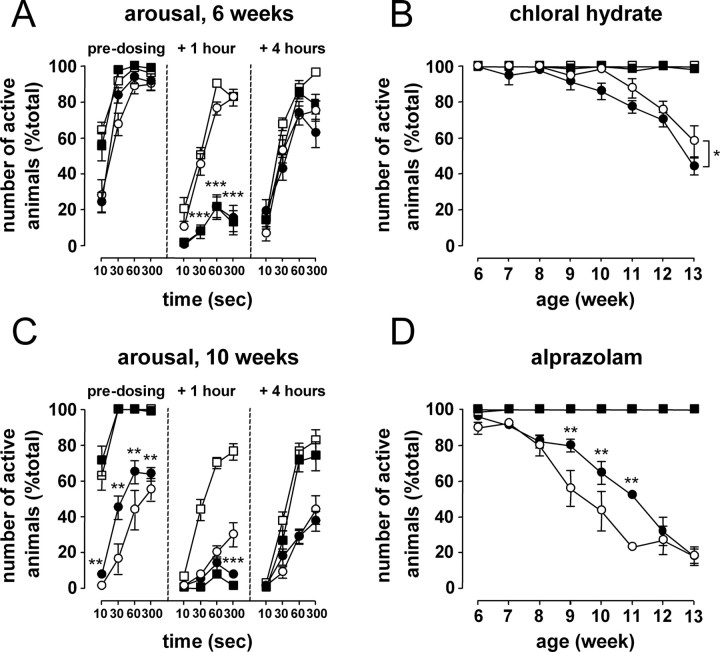
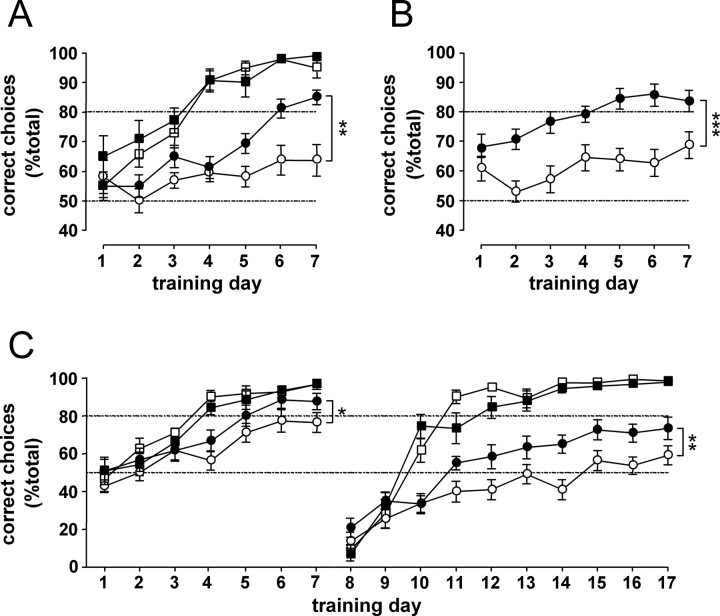
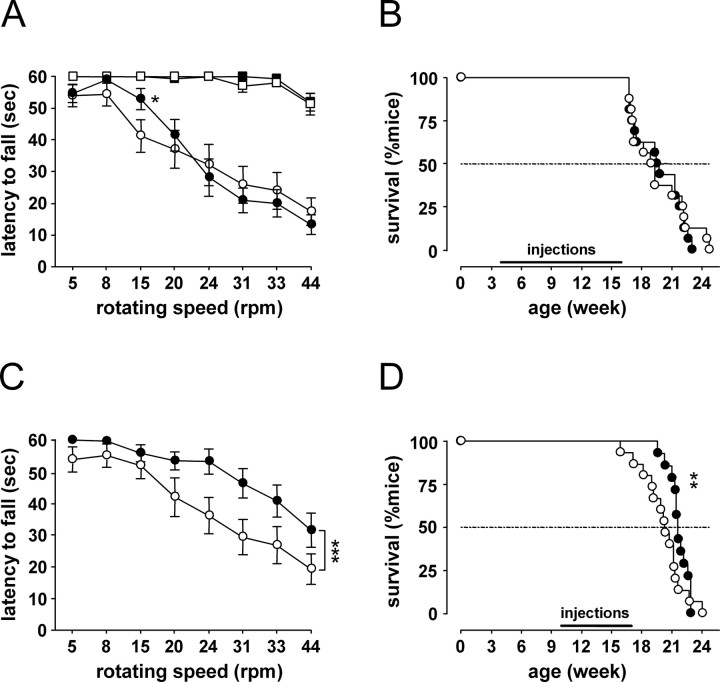
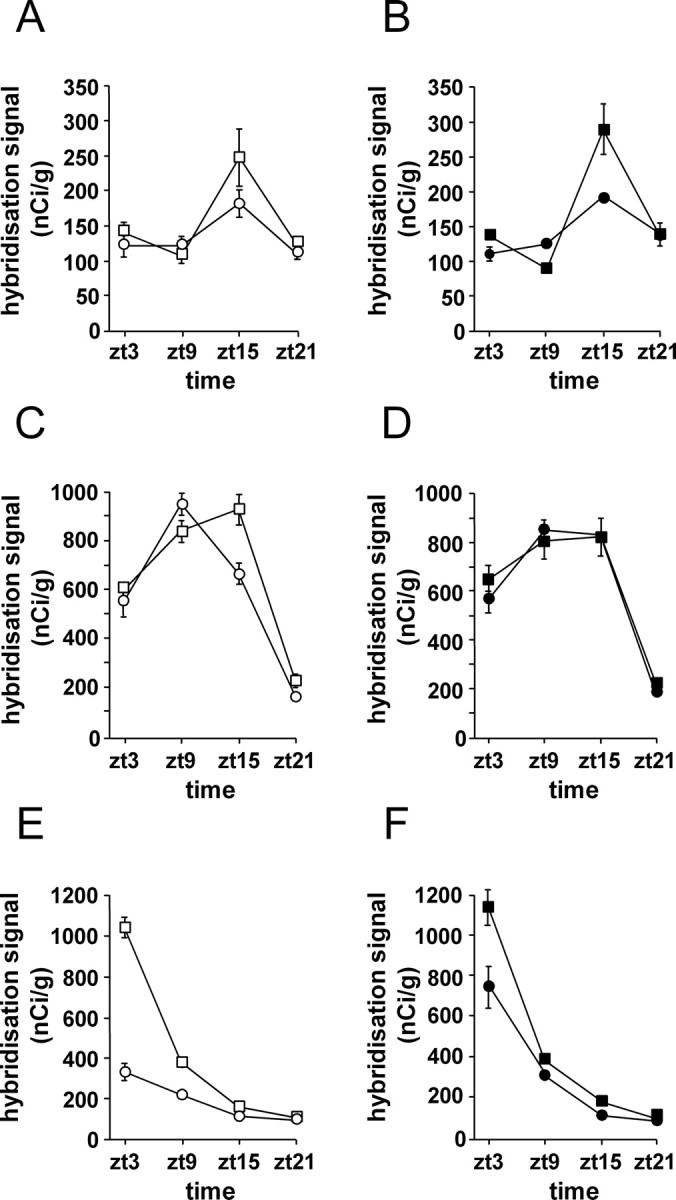
Transgenic R6/2 mice carrying the Huntington's disease (HD) mutation show disrupted circadian rhythms that worsen as the disease progresses. By 15 weeks of age, their abnormal circadian behavior mirrors that seen in HD patients and is accompanied by dysregulated clock gene expression in the circadian pacemaker, the suprachiasmatic nucleus (SCN). We found, however, that the electrophysiological output of the SCN assayed in vitro was normal. Furthermore, the endogenous rhythm of circadian gene expression, monitored in vitro by luciferase imaging of organotypical SCN slices removed from mice with disintegrated behavioral rhythms, was also normal. We concluded that abnormal behavioral and molecular circadian rhythms observed in R6/2 mice in vivo arise from dysfunction of brain circuitry afferent to the SCN, rather than from a primary deficiency within the pacemaker itself. Because circadian sleep disruption is deleterious to cognitive function, and cognitive decline is pronounced in R6/2 mice, we tested whether circadian and cognitive disturbances could be reversed by using a sedative drug to impose a daily cycle of sleep in R6/2 mice. Daily treatment with Alprazolam reversed the dysregulated expression of Per2 and also Prok2, an output factor of the SCN that controls behavioral rhythms. It also markedly improved cognitive performance of R6/2 mice in a two-choice visual discrimination task. Together, our data show for the first time that treatments aimed at restoring circadian rhythms may not only slow the cognitive decline that is such a devastating feature of HD but may also improve other circadian gene-regulated functions that are impaired in this disease.
Figures
References
-
- Bates GP, Harper PS, Jones L. Huntington's disease. Ed 3. Oxford: Oxford UP; 2002.
-
- Baudic S, Maison P, Dolbeau G, Boisse MF, Bartolomeo P, Dalla Barba G, Traykov L, Bachoud-Levi AC. Cognitive impairment related to apathy in early Huntington's disease. Dement Geriatr Cogn Disord. 2006;14:316–321. - PubMed
-
- Bianchetti A, Scuratti A, Zanetti O, Binetti G, Frisoni GB, Magni E, Trabucchi M. Predictors of mortality and institutionalization in Alzheimer disease patients 1 year after discharge from an Alzheimer dementia unit. Dementia. 1995;6:108–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases