

A molecular mechanism directly linking E-cadherin adhesion to initiation of epithelial cell surface polarity
- PMID: 17635938
- PMCID: PMC2064450
- DOI: 10.1083/jcb.200705094
A molecular mechanism directly linking E-cadherin adhesion to initiation of epithelial cell surface polarity
Abstract
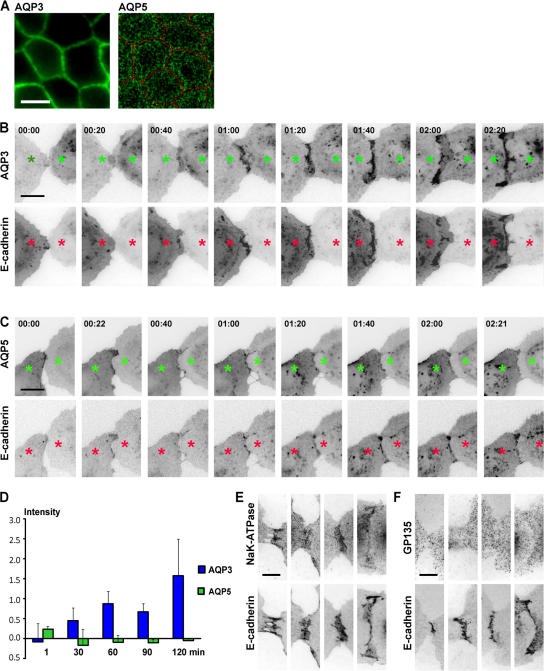
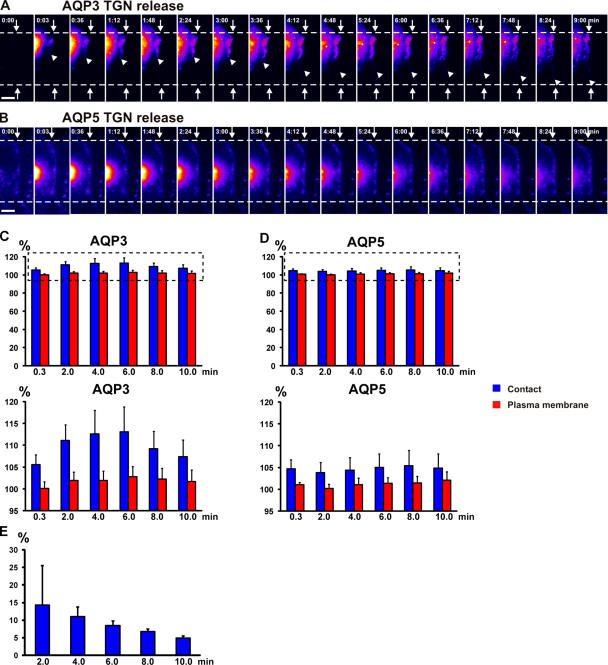
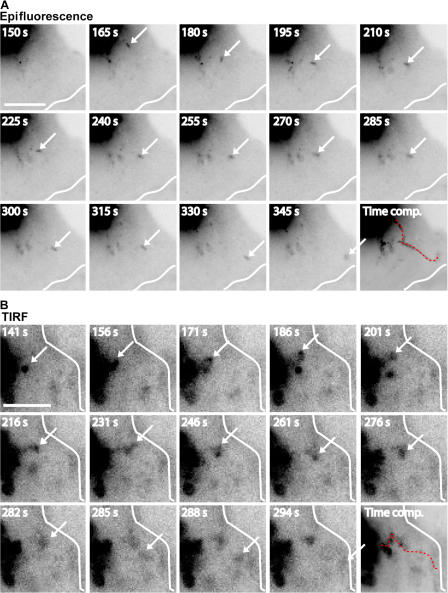
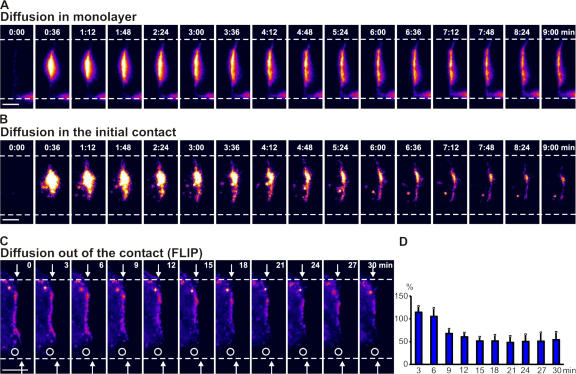
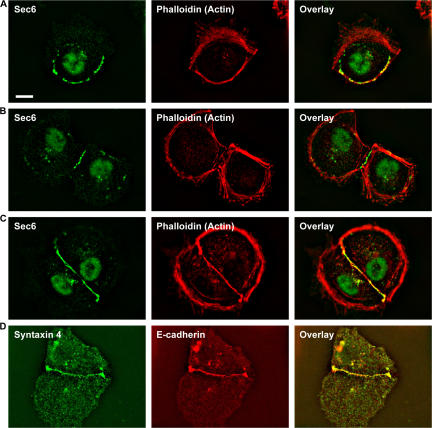
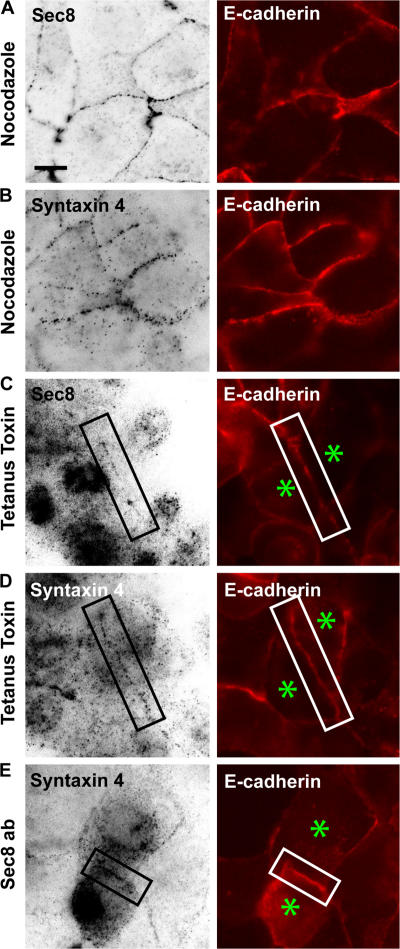
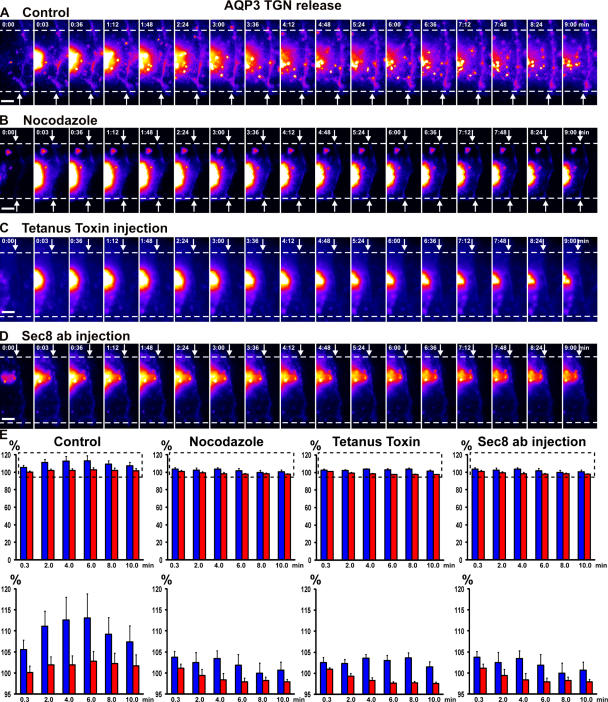
Mechanisms involved in maintaining plasma membrane domains in fully polarized epithelial cells are known, but when and how directed protein sorting and trafficking occur to initiate cell surface polarity are not. We tested whether establishment of the basolateral membrane domain and E-cadherin-mediated epithelial cell-cell adhesion are mechanistically linked. We show that the basolateral membrane aquaporin (AQP)-3, but not the equivalent apical membrane AQP5, is delivered in post-Golgi structures directly to forming cell-cell contacts where it co-accumulates precisely with E-cadherin. Functional disruption of individual components of a putative lateral targeting patch (e.g., microtubules, the exocyst, and soluble N-ethylmaleimide-sensitive factor attachment protein receptors) did not inhibit cell-cell adhesion or colocalization of the other components with E-cadherin, but each blocked AQP3 delivery to forming cell-cell contacts. Thus, components of the lateral targeting patch localize independently of each other to cell-cell contacts but collectively function as a holocomplex to specify basolateral vesicle delivery to nascent cell-cell contacts and immediately initiate cell surface polarity.
Figures
References
-
- Brunger, A.T. 2005. Structure and function of SNARE and SNARE-interacting proteins. Q. Rev. Biophys. 38:1–47. - PubMed
-
- Chang, J.T., V.R. Palanivel, I. Kinjyo, F. Schambach, A.M. Intlekofer, A. Banerjee, S.A. Longworth, K.E. Vinup, P. Mrass, J. Oliaro, et al. 2007. Asymmetric T lymphocyte division in the initiation of adaptive immune responses. Science. 315:1687–1691. - PubMed
