

IL-25 augments type 2 immune responses by enhancing the expansion and functions of TSLP-DC-activated Th2 memory cells
- PMID: 17635955
- PMCID: PMC2118667
- DOI: 10.1084/jem.20070406
IL-25 augments type 2 immune responses by enhancing the expansion and functions of TSLP-DC-activated Th2 memory cells
Abstract
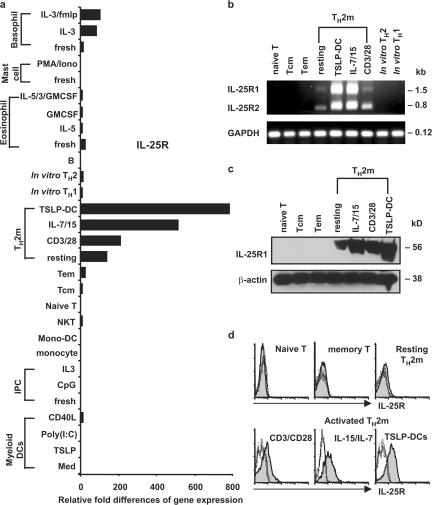
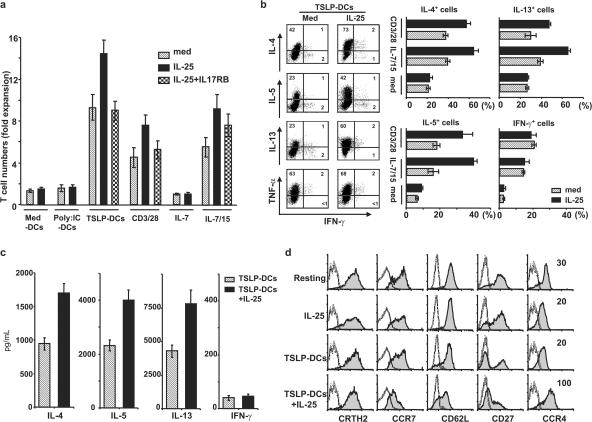
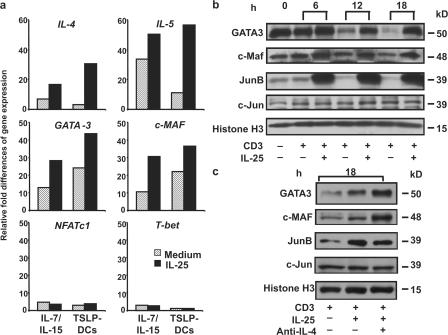
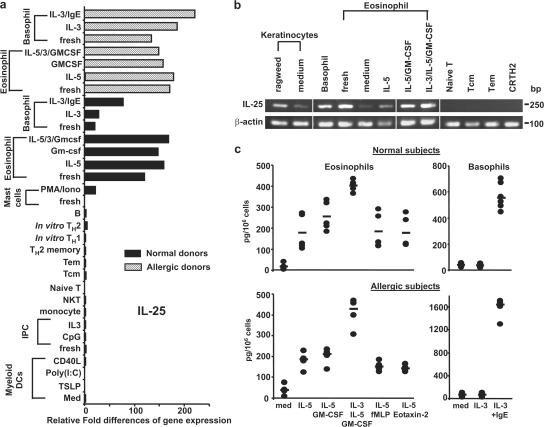
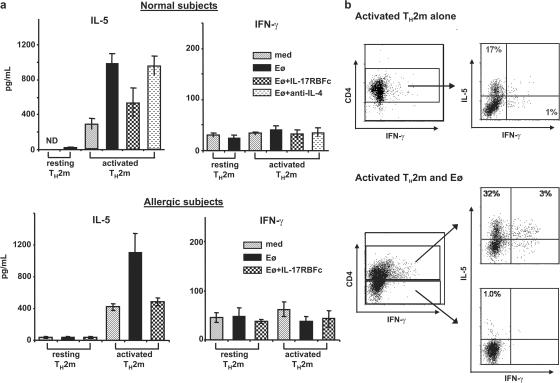
Interleukin (IL) 25 (IL-17E), a distinct member of the IL-17 cytokine family, plays important roles in evoking T helper type 2 (Th2) cell-mediated inflammation that features the infiltrations of eosinophils and Th2 memory cells. However, the cellular sources, target cells, and underlying mechanisms remain elusive in humans. We demonstrate that human Th2 memory cells expressing distinctive levels of IL-25 receptor (R) are one of the responding cell types. IL-25 promotes cell expansion and Th2 cytokine production when Th2 central memory cells are stimulated with thymic stromal lymphopoietin (TSLP)-activated dendritic cells (DCs), homeostatic cytokines, or T cell receptor for antigen triggering. The enhanced functions of Th2 memory cells induced by IL-25 are associated with sustained expression of GATA-3, c-MAF, and JunB in an IL-4-independent manner. Although keratinocytes, mast cells, eosinophils, and basophils express IL-25 transcripts, activated eosinophils and basophils from normal and atopic subjects were found to secrete bioactive IL-25 protein, which augments the functions of Th2 memory cells. Elevated expression of IL-25 and IL-25R transcripts was observed in asthmatic lung tissues and atopic dermatitis skin lesions, linking their possible roles with exacerbated allergic disorders. Our results provide a plausible explanation that IL-25 produced by innate effector eosinophils and basophils may augment the allergic inflammation by enhancing the maintenance and functions of adaptive Th2 memory cells.
Figures


References
-
- Cohn, L., J.A. Elias, and G.L. Chupp. 2004. Asthma: mechanisms of disease persistence and progression. Annu. Rev. Immunol. 22:789–815. - PubMed
-
- Wills-Karp, M. 1999. Immunologic basis of antigen-induced airway hyper responsiveness. Annu. Rev. Immunol. 17:255–281. - PubMed
-
- Gleich, G.J., C.R. Adolphson, and K.M. Leiferman. 1993. The biology of the eosinophilic leukocyte. Annu. Rev. Med. 44:85–101. - PubMed
-
- Rothenberg, M.E. 1998. Eosinophilia. N. Engl. J. Med. 338:1592–1600. - PubMed
-
- Pan, G., D. French, W. Mao, M. Maruoka, P. Risser, J. Lee, J. Foster, S. Aggarwal, K. Nicholes, S. Guillet, et al. 2001. Forced expression of murine IL-17E induces growth retardation, jaundice, a Th2-biased response, and multiorgan inflammation in mice. J. Immunol. 167:6559–6567. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
