

The mitochondrial respiratory chain controls intracellular calcium signaling and NFAT activity essential for heart formation in Xenopus laevis
- PMID: 17636012
- PMCID: PMC2099623
- DOI: 10.1128/MCB.01946-06
The mitochondrial respiratory chain controls intracellular calcium signaling and NFAT activity essential for heart formation in Xenopus laevis
Abstract
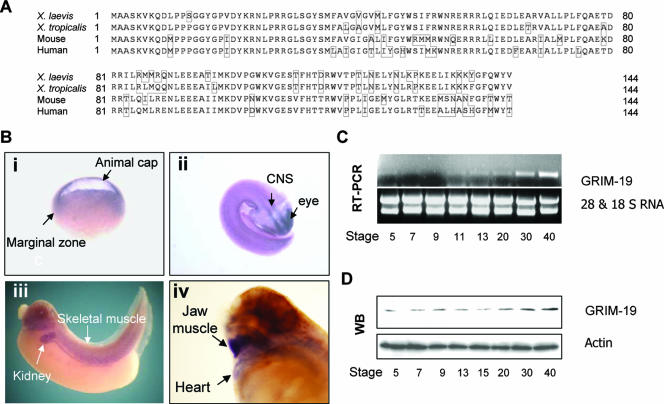
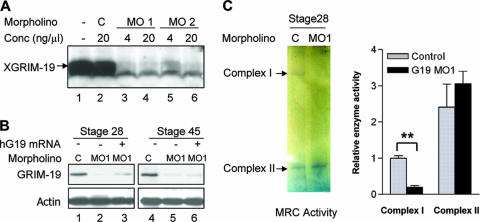
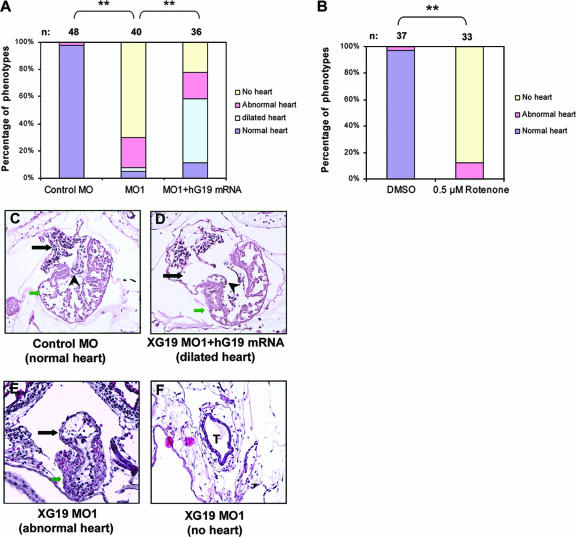
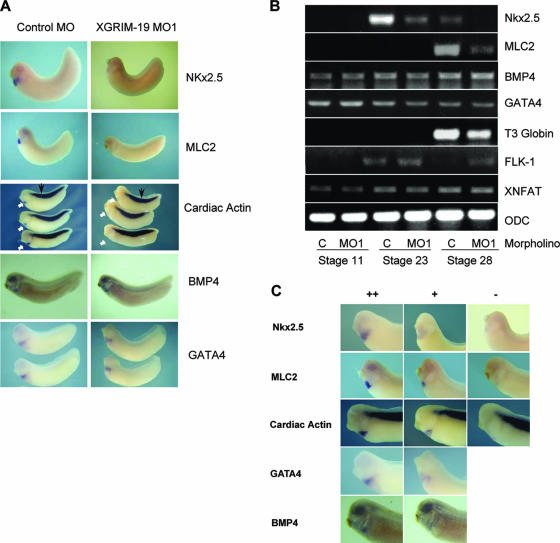
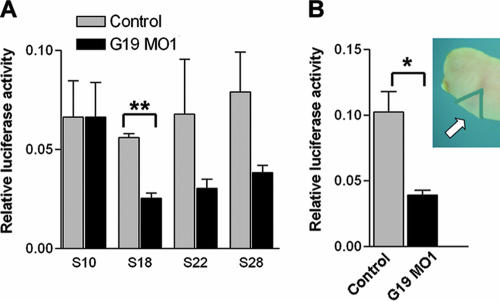
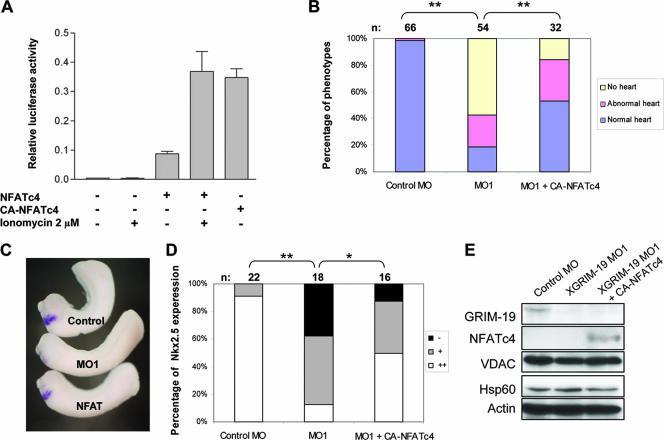
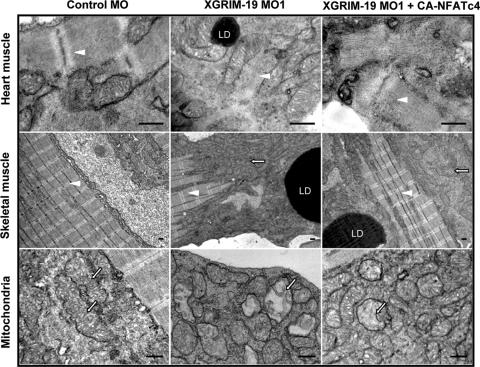
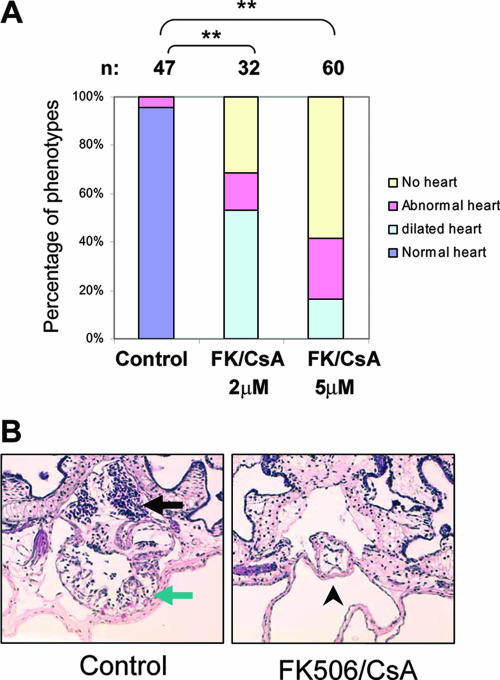
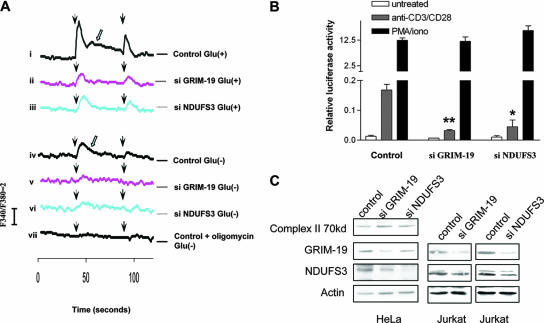
The mitochondrial respiratory chain (MRC) plays crucial roles in cellular energy production. However, its function in early embryonic development remains largely unknown. To address this issue, GRIM-19, a newly identified MRC complex I subunit, was knocked down in Xenopus laevis embryos. A severe deficiency in heart formation was observed, and the deficiency could be rescued by reintroducing human GRIM-19 mRNA. The mechanism involved was further investigated. We found that the activity of NFAT, a transcription factor family that contributes to early organ development, was downregulated in GRIM-19 knockdown embryos. Furthermore, the expression of a constitutively active form of mouse NFATc4 in these embryos rescued the heart developmental defects. NFAT activity is controlled by a calcium-dependent protein phosphatase, calcineurin, which suggests that calcium signaling may be disrupted by GRIM-19 knockdown. Indeed, both the calcium response and calcium-induced NFAT activity were impaired in the GRIM-19 or NDUFS3 (another complex I subunit) knockdown cell lines. We also showed that NFAT can rescue expression of Nkx2.5, which is one of the key genes for early heart development. Our data demonstrated the essential role of MRC in heart formation and revealed the signal transduction and gene expression cascade involved in this process.
Figures
Similar articles
-
Developmental regulation of the mouse IGF-I exon 1 promoter region by calcineurin activation of NFAT in skeletal muscle.Am J Physiol Cell Physiol. 2007 May;292(5):C1887-94. doi: 10.1152/ajpcell.00506.2006. Epub 2007 Jan 17. Am J Physiol Cell Physiol. 2007. PMID: 17229811
-
NFAT directly regulates Nkx2-5 transcription during cardiac cell differentiation.Biol Cell. 2009 Jun;101(6):335-49. doi: 10.1042/BC20080108. Biol Cell. 2009. PMID: 18828760
-
Dosage-dependent transcriptional regulation by the calcineurin/NFAT signaling in developing myocardium transition.Dev Biol. 2007 Mar 15;303(2):825-37. doi: 10.1016/j.ydbio.2006.11.036. Epub 2006 Dec 2. Dev Biol. 2007. PMID: 17198697
-
Calcineurin activates interleukin-6 transcription in mouse skeletal muscle in vivo and in C2C12 myotubes in vitro.Am J Physiol Regul Integr Comp Physiol. 2010 Jan;298(1):R198-210. doi: 10.1152/ajpregu.00325.2009. Epub 2009 Nov 11. Am J Physiol Regul Integr Comp Physiol. 2010. PMID: 19907005 Free PMC article.
-
Expression and function of Xenopus laevis p75(NTR) suggest evolution of developmental regulatory mechanisms.J Neurobiol. 2001 Nov 5;49(2):79-98. doi: 10.1002/neu.1067. J Neurobiol. 2001. PMID: 11598917 Review.
Cited by
-
Mitochondrial Ca2+ cycling facilitates activation of the transcription factor NFAT in sensory neurons.J Neurosci. 2009 Sep 30;29(39):12101-14. doi: 10.1523/JNEUROSCI.3384-09.2009. J Neurosci. 2009. PMID: 19793968 Free PMC article.
-
Lessons from the lily pad: Using Xenopus to understand heart disease.Drug Discov Today Dis Models. 2008 Fall;5(3):141-146. doi: 10.1016/j.ddmod.2009.02.006. Drug Discov Today Dis Models. 2008. PMID: 19802378 Free PMC article.
-
Increased COX-1 expression in benign prostate epithelial cells is triggered by mitochondrial dysfunction.Am J Clin Exp Urol. 2022 Aug 15;10(4):234-245. eCollection 2022. Am J Clin Exp Urol. 2022. PMID: 36051613 Free PMC article.
-
GRIM-19 is essential for maintenance of mitochondrial membrane potential.Mol Biol Cell. 2008 May;19(5):1893-902. doi: 10.1091/mbc.e07-07-0683. Epub 2008 Feb 20. Mol Biol Cell. 2008. PMID: 18287540 Free PMC article.
-
DNA methylation is developmentally regulated for genes essential for cardiogenesis.J Am Heart Assoc. 2014 Jun 19;3(3):e000976. doi: 10.1161/JAHA.114.000976. J Am Heart Assoc. 2014. PMID: 24947998 Free PMC article.
References
-
- Angell, J. E., D. J. Lindner, P. S. Shapiro, E. R. Hofmann, and D. V. Kalvakolanu. 2000. Identification of GRIM-19, a novel cell death-regulatory gene induced by the interferon-beta and retinoic acid combination, using a genetic approach. J. Biol. Chem. 275:33416-33426. - PubMed
-
- Berridge, M. J., M. D. Bootman, and H. L. Roderick. 2003. Calcium signalling: dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 4:517-529. - PubMed
-
- Bezprozvanny, I., and B. E. Ehrlich. 1993. ATP modulates the function of inositol 1,4,5-trisphosphate-gated channels at two sites. Neuron 10:1175-1184. - PubMed
-
- Bushdid, P. B., H. Osinska, R. R. Waclaw, J. D. Molkentin, and K. E. Yutzey. 2003. NFATc3 and NFATc4 are required for cardiac development and mitochondrial function. Circ. Res. 92:1305-1313. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous