

Epigenetic regulation of protein-coding and microRNA genes by the Gfi1-interacting tumor suppressor PRDM5
- PMID: 17636019
- PMCID: PMC2099216
- DOI: 10.1128/MCB.00762-07
Epigenetic regulation of protein-coding and microRNA genes by the Gfi1-interacting tumor suppressor PRDM5
Abstract
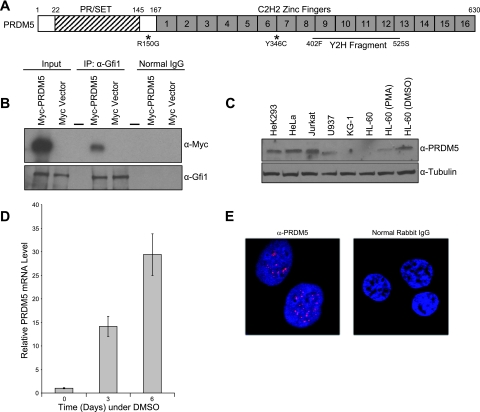
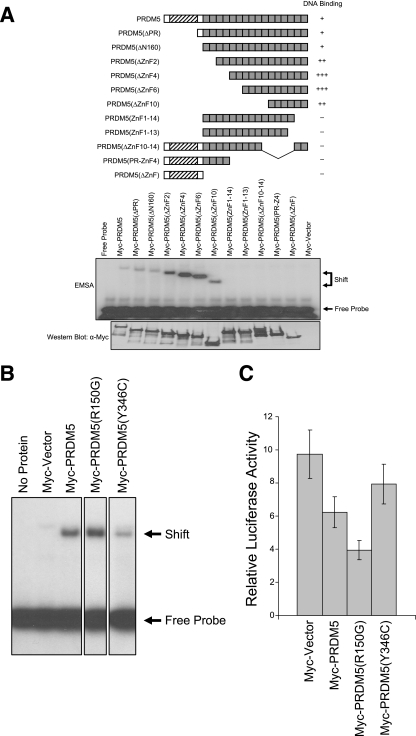
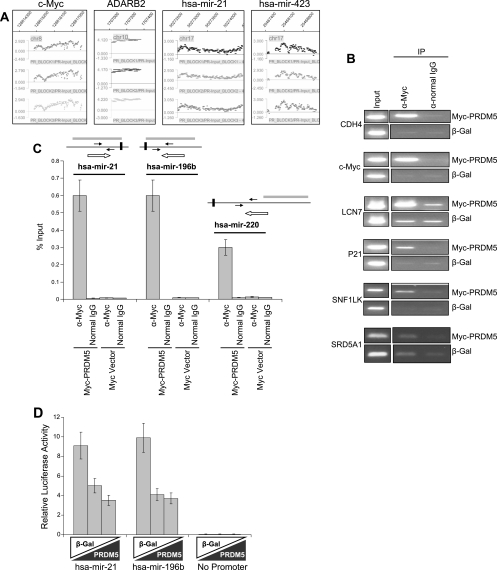
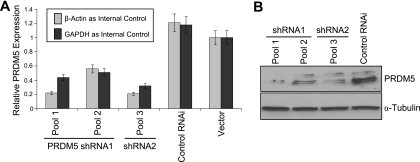
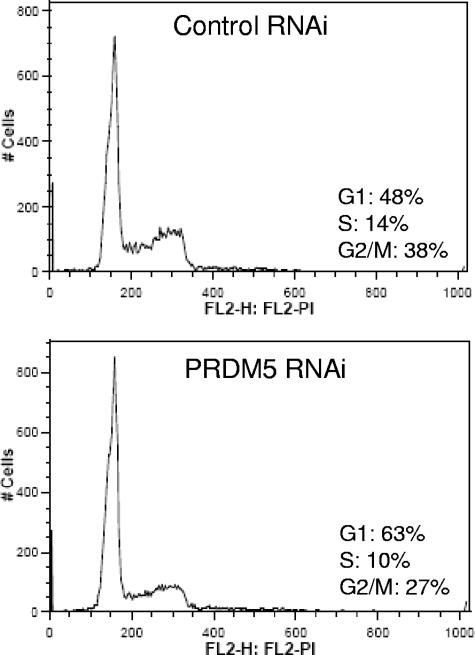
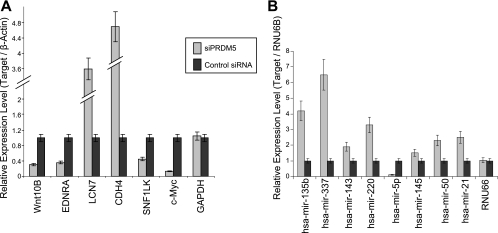
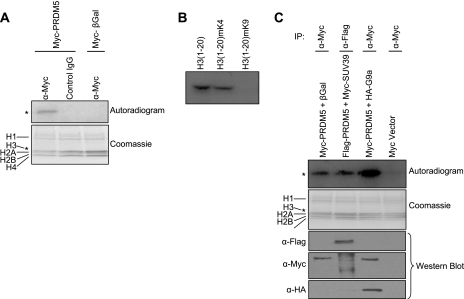
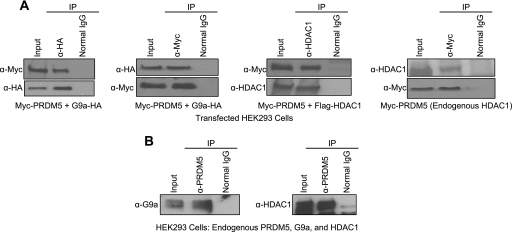
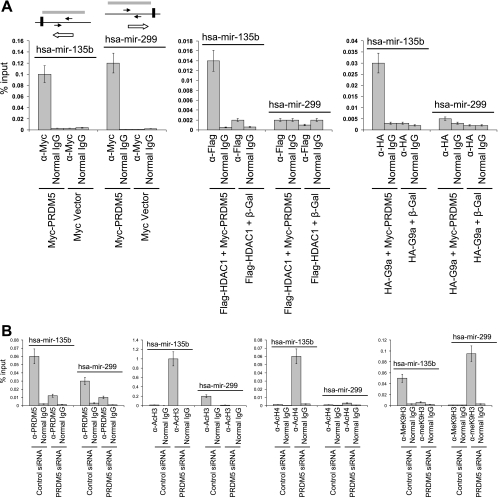
Gfi1 transcriptionally governs hematopoiesis, and its mutations produce neutropenia. In an effort to identify Gfi1-interacting proteins and also to generate new candidate genes causing neutropenia, we performed a yeast two-hybrid screen with Gfi1. Among other Gfi1-interacting proteins, we identified a previously uncharacterized member of the PR domain-containing family of tumor suppressors, PRDM5. PRDM5 has 16 zinc fingers, and we show that it acts as a sequence-specific, DNA binding transcription factor that targets hematopoiesis-associated protein-coding and microRNA genes, including many that are also targets of Gfi1. PRDM5 epigenetically regulates transcription similarly to Gfi1: it recruits the histone methyltransferase G9a and class I histone deacetylases to its target gene promoters and demonstrates repressor activity on synthetic reporters; on endogenous target genes, however, it functions as an activator, in addition to a repressor. Interestingly, genes that PRDM5 activates, as opposed to those it represses, are also targets of Gfi1, suggesting a competitive mechanism through which two repressors could cooperate in order to become transcriptional activators. In neutropenic patients, we identified PRDM5 protein sequence variants perturbing transcriptional function, suggesting a potentially important role in hematopoiesis.
Figures
References
-
- Abuin, A., G. M. Hansen, and B. Zambrowicz. 2007. Gene trap mutagenesis. Handb. Exp. Pharmacol. 178:129-147. - PubMed
-
- Ancelin, K., U. C. Lange, P. Hajkova, R. Schneider, A. J. Bannister, T. Kouzarides, and M. A. Surani. 2006. Blimp1 associates with Prmt5 and directs histone arginine methylation in mouse germ cells. Nat. Cell Biol. 8:623-630. - PubMed
-
- Canote, R., Y. Du, T. Carling, F. Tian, Z. Peng, and S. Huang. 2002. The tumor suppressor gene RIZ in cancer gene therapy (review). Oncol. Rep. 9:57-60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous