

A viral adaptor protein modulating casein kinase II activity induces cytopathic effects in permissive cells
- PMID: 17636126
- PMCID: PMC1920537
- DOI: 10.1073/pnas.0705533104
A viral adaptor protein modulating casein kinase II activity induces cytopathic effects in permissive cells
Abstract
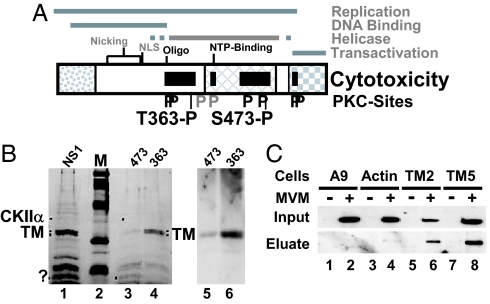
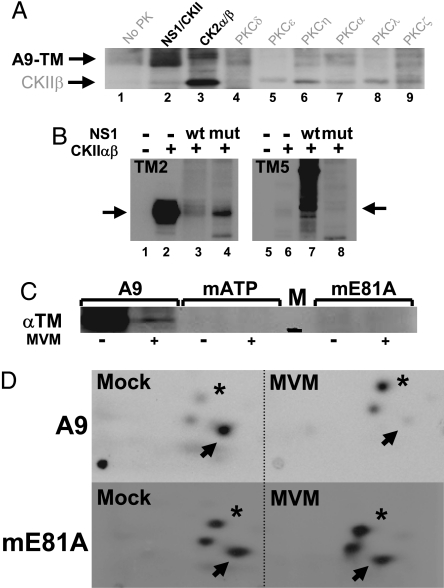
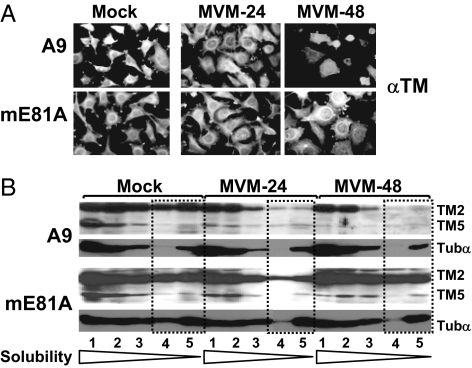
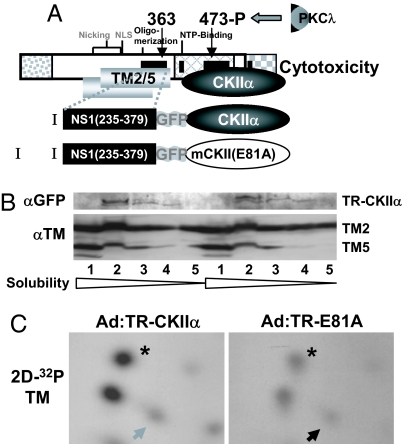
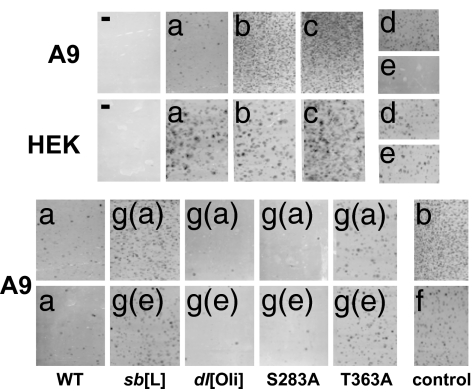
Autonomous parvoviruses induce severe morphological and physiological alterations in permissive host cells, eventually leading to cell lysis and release of progeny virions. Viral cytopathic effects (CPE) result from specific rearrangements and destruction of cytoskeletal micro- and intermediate filaments. We recently reported that inhibition of endogenous casein kinase II (CKII) protects target cells from parvovirus minute virus of mice (MVM)-induced CPE, pointing to this kinase as an effector of MVM toxicity. The present work shows that the parvoviral NS1 protein mediates CKII-dependent cytoskeletal alterations and cell death. NS1 can act as an adaptor molecule, linking the cellular protein kinase CKIIalpha to tropomyosin and thus modulating the substrate specificity of the kinase. This action results in an altered tropomyosin phosphorylation pattern both in vitro and in living cells. The capacity of NS1 to induce CPE was impaired by mutations abolishing binding with either CKIIalpha or tropomyosin. The cytotoxic adaptor function of NS1 was confirmed with fusion peptides, where the tropomyosin-binding domain of NS1 and CKIIalpha are physically linked. These adaptor peptides were able to mimic NS1 in its ability to induce death of transformed MVM-permissive cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Geletneky K, Herrero YCM, Rommelaere J, Schlehofer JR. J Vet Med. 2005;52:327–330. - PubMed
-
- Nüesch JP. In: Parvoviruses. Kerr JR, Cotmore S, Bloom ME, Linden RM, Parrish CR, editors. London: Edward Arnold; 2006. pp. 275–290.
-
- Corbau R, Duverger V, Rommelaere J, Nüesch JP. Virology. 2000;278:151–167. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
