

Avian circannual clocks: adaptive significance and possible involvement of energy turnover in their proximate control
- PMID: 17638688
- PMCID: PMC2606758
- DOI: 10.1098/rstb.2007.2147
Avian circannual clocks: adaptive significance and possible involvement of energy turnover in their proximate control
Abstract
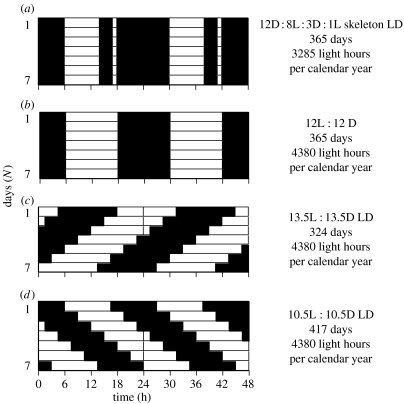
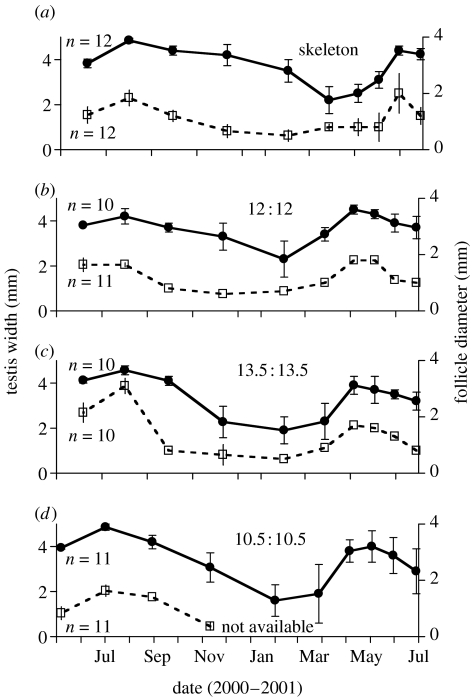
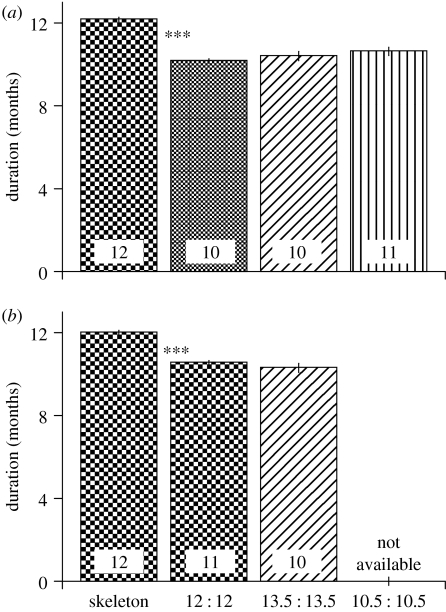
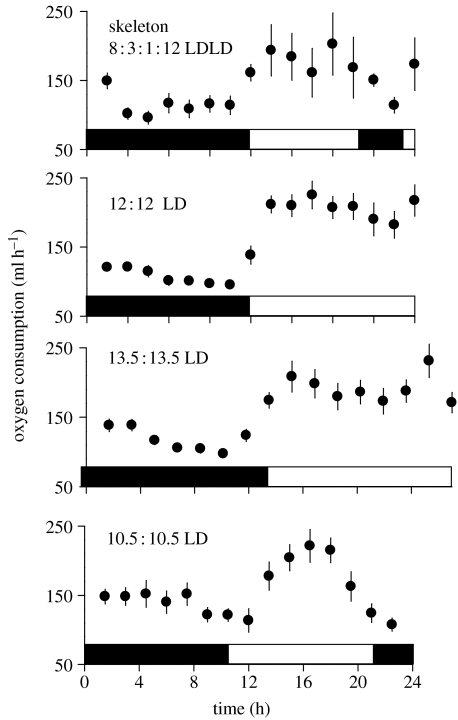
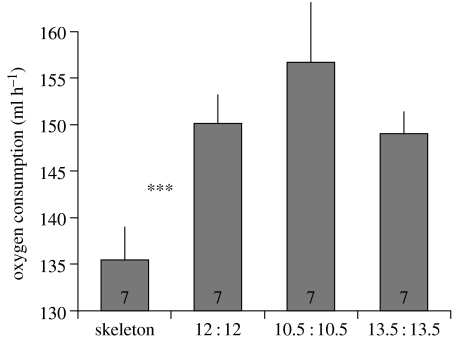
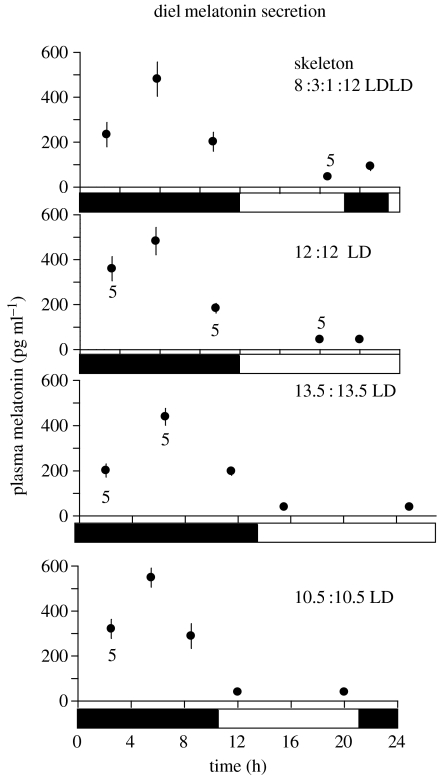
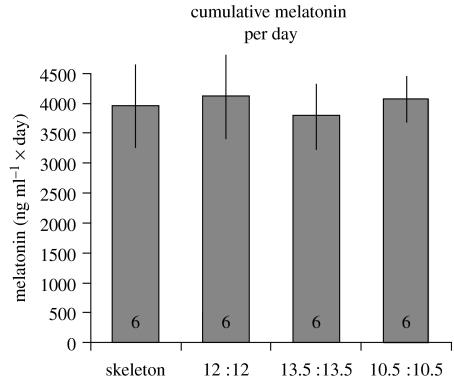
Endogenous circannual clocks are found in many long-lived organisms, but are best studied in mammal and bird species. Circannual clocks are synchronized with the environment by changes in photoperiod, light intensity and possibly temperature and seasonal rainfall patterns. Annual timing mechanisms are presumed to have important ultimate functions in seasonally regulating reproduction, moult, hibernation, migration, body weight and fat deposition/stores. Birds that live in habitats where environmental cues such as photoperiod are poor predictors of seasons (e.g. equatorial residents, migrants to equatorial/tropical latitudes) rely more on their endogenous clocks than birds living in environments that show a tight correlation between photoperiod and seasonal events. Such population-specific/interspecific variation in reliance on endogenous clocks may indicate that annual timing mechanisms are adaptive. However, despite the apparent adaptive importance of circannual clocks, (i) what specific adaptive value they have in the wild and (ii) how they function are still largely untested. Whereas circadian clocks are hypothesized to be generated by molecular feedback loops, it has been suggested that circannual clocks are either based upon (i) a de-multiplication ('counting') of circadian days, (ii) a sequence of interdependent physiological states, or (iii) one or more endogenous oscillators, similar to circadian rhythms. We tested the de-multiplication of days (i) versus endogenous regulation hypotheses (ii) and (iii) in captive male and female house sparrows (Passer domesticus). We assessed the period of reproductive (testicular and follicular) cycles in four groups of birds kept either under photoperiods of LD 12L:12D (period length: 24h), 13.5L:13.5D (27 h), 10.5L:10.5D (23 h) or 12D:8L:3D:1L (24-h skeleton photoperiod), respectively, for 15 months. Contrary to predictions from the de-multiplication hypothesis, individuals experiencing 27-h days did not differ (i.e. did not have longer) annual reproductive rhythms than individuals from the 21- or 24-h day groups. However, in line with predictions from endogenous regulation, birds in the skeleton group had significantly longer circannual period lengths than all other groups. Birds exposed to skeleton photoperiods experienced fewer light hours per year than all other groups (3285 versus 4380) and had a lower daily energy expenditure, as tested during one point of the annual cycle using respirometry. Although our results are tantalizing, they are still preliminary as birds were only studied over a period of 15 months. Nevertheless, the present data fail to support a 'counting of circadian days' and instead support hypotheses proposing whole-organism processes as the mechanistic basis for circannual rhythms. We propose a novel energy turnover hypothesis which predicts a dependence of the speed of the circannual clock on the overall energy expenditure of an organism.
Figures
Similar articles
-
Circannual testis and moult cycles persist under photoperiods that disrupt circadian activity and clock gene cycles in spotted munia.J Exp Biol. 2017 Nov 15;220(Pt 22):4162-4168. doi: 10.1242/jeb.167809. Epub 2017 Sep 15. J Exp Biol. 2017. PMID: 28916681
-
Seasonality in a temperate zone bird can be entrained by near equatorial photoperiods.Proc Biol Sci. 2007 Mar 7;274(1610):721-5. doi: 10.1098/rspb.2006.0067. Proc Biol Sci. 2007. PMID: 17254997 Free PMC article.
-
Circannual rhythms in birds.Curr Opin Neurobiol. 2003 Dec;13(6):770-8. doi: 10.1016/j.conb.2003.10.010. Curr Opin Neurobiol. 2003. PMID: 14662381 Review.
-
Avian circannual systems: persistence and sex differences.Gen Comp Endocrinol. 2013 Sep 1;190:61-7. doi: 10.1016/j.ygcen.2013.04.013. Epub 2013 Apr 28. Gen Comp Endocrinol. 2013. PMID: 23631901 Review.
-
Seasonal Time Keeping in a Long-Distance Migrating Shorebird.J Biol Rhythms. 2016 Oct;31(5):509-21. doi: 10.1177/0748730416655929. Epub 2016 Jul 27. J Biol Rhythms. 2016. PMID: 27466352
Cited by
-
Night Migratory Songbirds Exhibit Metabolic Ability to Support High Aerobic Capacity during Migration.ACS Omega. 2020 Oct 22;5(43):28088-28095. doi: 10.1021/acsomega.0c03691. eCollection 2020 Nov 3. ACS Omega. 2020. PMID: 33163791 Free PMC article.
-
Optimal annual routines: behaviour in the context of physiology and ecology.Philos Trans R Soc Lond B Biol Sci. 2008 Jan 27;363(1490):301-19. doi: 10.1098/rstb.2007.2141. Philos Trans R Soc Lond B Biol Sci. 2008. PMID: 17673417 Free PMC article.
-
Artificial light at night advances avian reproductive physiology.Proc Biol Sci. 2013 Feb 13;280(1756):20123017. doi: 10.1098/rspb.2012.3017. Print 2013 Apr 7. Proc Biol Sci. 2013. PMID: 23407836 Free PMC article.
-
Nocturnal One-Hour Lighting Stimulates Gonadal Development and Lowers Fat Deposition in Male Mule Ducks.Animals (Basel). 2021 Feb 26;11(3):614. doi: 10.3390/ani11030614. Animals (Basel). 2021. PMID: 33652787 Free PMC article.
-
Hormonally mediated effects of artificial light at night on behavior and fitness: linking endocrine mechanisms with function.J Exp Biol. 2018 Mar 15;221(Pt 6):jeb156893. doi: 10.1242/jeb.156893. J Exp Biol. 2018. PMID: 29545373 Free PMC article. Review.
References
-
- Aschoff J. Tierische Periodik unter dem Einfluß von Zeitgebern. Zeitschrift für Tierpsychologie. 1958;15:1–30.
-
- Aschoff J. Circadian rhythms: Influences of internal and external factors on the period measured in constant conditions. Zeitschrift für Tierpsychologie. 1979;49:225–249. - PubMed
-
- Bairlein F, Gwinner E. Nutritional mechanisms and temporal control of migratory energy accumulation in birds. Annu. Rev. Nutr. 1994;14:187–215. doi:10.1146/annurev.nu.14.070194.001155 - DOI - PubMed
-
- Barta Z, McNamara J.M, Houston A.I, Weber T.P, Hedenstro¨m A, Feró O. Optimal moult strategies in migratory birds. Phil. Trans. R. Soc. B. 2008;363:211–229. doi:10.1098/rstb.2007.2136 - DOI - PMC - PubMed
-
- Bentley G.E, Goldsmith A.R, Dawson A, Briggs C, Pemberton M. Decreased light intensity alters the perception of daylength by male European starlings (Sturnus vulgaris) J. Biol. Rhythms. 1998;13:140–150. doi:10.1177/074873098128999998 - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
