

doi: 10.1101/gad.1549407.
Oncogenic Ras-induced secretion of IL6 is required for tumorigenesis
Affiliations
- PMID: 17639077
- PMCID: PMC1920165
- DOI: 10.1101/gad.1549407
Item in Clipboard
Oncogenic Ras-induced secretion of IL6 is required for tumorigenesis
Genes Dev.
.
Abstract
Ras is mutated to remain in the active oncogenic state in many cancers. As Ras has proven difficult to target therapeutically, we searched for secreted, druggable proteins induced by Ras that are required for tumorigenesis. We found that Ras induces the secretion of cytokine IL6 in different cell types, and that knockdown of IL6, genetic ablation of the IL6 gene, or treatment with a neutralizing IL6 antibody retard Ras-driven tumorigenesis. IL6 appears to act in a paracrine fashion to promote angiogenesis and tumor growth. Inhibiting IL6 may therefore have therapeutic utility for treatment of cancers characterized by oncogenic Ras mutations.
Figures
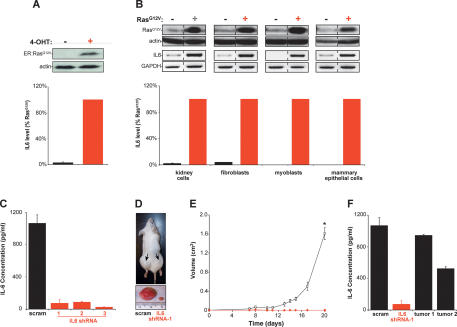
Oncogenic Ras-induced secretion of IL6 is required for Ras-mediated tumorigenesis. (A) IL6-secreted protein levels increase upon activation of Ras. Human kidney cells expressing T/t-Ag, hTERT, and ER:RasG12V (and p110-CAAX) were confirmed by immunoblot to up-regulate ER:RasG12V in the presence of 4-OHT (top), which led to elevated levels of IL6 (percent relative to RasG12V-expressing cells) present in cell medium as detected by ELISA (bottom). (Actin) Loading control. (B) Oncogenic Ras increases secreted IL6 independent of cell type. The indicated four primary human cell types expressing T/t-Ag and hTERT and, where indicated, also RasG12V, as assessed by immunoblot (top), were shown to increase the levels of IL6 mRNA, as assessed by RT–PCR (middle), and secreted IL6 protein (percent relative to RasG12V-expressing cells), as assessed by ELISA (bottom) upon expression of RasG12V. (Actin) Loading control; (GAPDH) RT–PCR control. (C) Primary human kidney cells expressing T/t-Ags, hTERT, RasG12V, and shRNA-1, shRNA-2, and shRNA-3, but not a scramble (scram) control sequence, exhibited a decrease in secreted IL6 as assessed by ELISA. (D) Knockdown of IL6 (IL6 shRNA-1) in the aforementioned human kidney cells induced decreased tumor growth, as visualized in a representative mouse (arrow, tumor) or ressected tumors, when compared with scramble control-treated cells (scram). (E) IL6 is required for Ras-driven tumor growth. A plot of tumor volume (cubic centimeters) ± standard error versus time in days for the aforementioned human kidney cells expressing the scramble control sequence (open box) or the three different IL6 shRNAs (as growth kinetics were identical, tumor growth in mice injected with cells expressing any of the three IL6 shRNAs is shown with one symbol [red diamond]) when injected in mice. (*) P < 0.001 (F) Two tumors (tumor 1, tumor 2), eventually arising when the aforementioned human kidney cells expressing IL6 shRNA-1 were isolated, were cultured briefly in hygromycin to enrich for tumor cells and assayed for secreted IL6 by ELISA, with the finding that these tumors exhibited increased IL6 compared with the original IL6 shRNA-1 donor cells, almost to the level of scramble (scram) control cells.
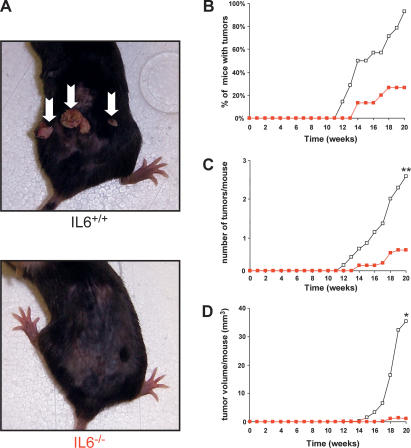
IL6−/− mice are resistant to carcinogen-induced skin tumors. Fourteen IL6+/+ and 15 IL6−/− mice were treated with a single topical application of DMBA, followed 1 wk later by twice-weekly topical applications of TPA for 20 wk to induce skin tumors. (A) Reduction of spontaneous tumors in IL6−/− mice. Representative mice of the indicated genotype at 20 wk. (Arrows) Tumors. (B) Reduction in the number of IL6−/− mice having tumors. Percentage of IL6+/+ (open box) and IL6−/− (red box) mice with tumors versus time after initial application of DMBA (weeks). (C) Reduction in the number of tumors in IL6−/− mice. Mean number of tumors per IL6+/+ (open box) and IL6−/− (red box) mouse versus time after initial application of DMBA (weeks). (**) P < 0.01 (D) Reduction in tumor volume in IL6−/− mice. Mean tumor volume per IL6+/+ (open box) and IL6−/− (red box) mouse versus time after initial application of DMBA (weeks). (*) P < 0.05.
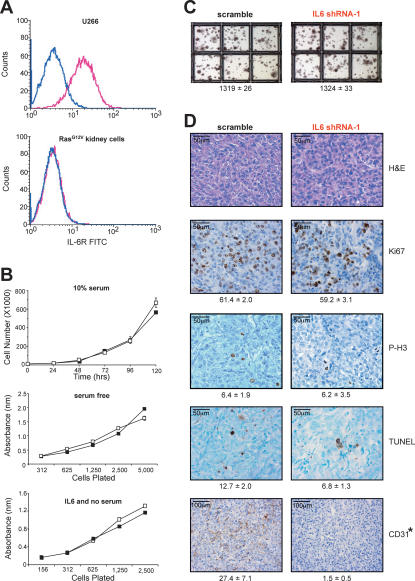
IL6 acts in a paracrine manner to promote angiogenesis. (A) Detection of IL6R (pink profile) by flow-cytometric analysis in positive control U266 cells (top graph), but not RasG12V-transformed human kidney cells (bottom graph). No antibody (blue profile) serves as a negative control. (B) IL6 does not alter the growth of RasG12V-transformed human kidney cells. Absorbance ± standard error versus time (days) as measured by the MTT assay of RasG12V-transformed human kidney cells stably expressing IL6 shRNA-1 (black box) or the appropriate scramble sequence (open box) cultured in 10% serum (top) or serum-free (middle) medium, or the same kidney cells lacking RasG12V plated at the indicated densities in serum-free medium (black box) or serum-free medium containing 100 pg/mL IL6 (open box). (C) Similar anchorage-independent growth of RasG12V-transformed human kidney cells stably expressing IL6 shRNA-1 versus the appropriate scramble sequence. Average number of colonies ± standard error calculated from three independent experiments conducted in triplicate. (D) Loss of IL6 inhibits angiogenesis. Tumors from RasG12V-transformed human kidney cells stably expressing IL6 shRNA-1 or the appropriate scramble sequence were excised, formalin-fixed, and stained for H&E or assayed for Ki67, phospho-histone H3 (P-H3), TUNEL, or CD31-positive (dark brown) cells. (Bottom) Average number of marker-positive cells ± standard deviation from five independent fields of two to four different tumors. (*) P < 0.01)
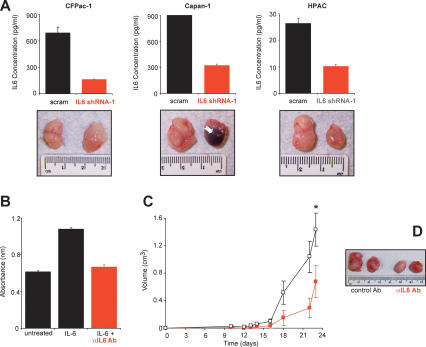
IL6 as a therapeutic target. (A) Knockdown of IL6 inhibits tumor growth. (Top panel) Reduction of IL6, as detected by ELISA from the conditioned medium of the indicated three human pancreatic cancer cell lines expressing IL6 shRNA-1, but not the scramble (scram) control counterpart. (Bottom panel) Representative tumors from the indicated three human pancreatic cancer cell lines stably expressing an IL6 shRNA-1 or scramble sequence after injection subcutaneously into the flanks of mice. (Arrow) Region of necrosis. (B) Neutralization of IL6 activity by an anti-IL6 antibody (αIL6 Ab). Cell numbers as measured by average absorbance ± standard error using the MTT assay 48 h after B9 cells were plated in serum-free medium (untreated), in the presence of IL6 to stimulate cell proliferation (IL6) or in the presence of IL6 and αIL6 Ab (IL6 + αIL6 Ab). (C) An IL6-neutralizing antibody inhibits Ras-driven tumor growth. Tumor volume (cubic centimeters) ± standard error versus time (days) of RasG12V-transformed human kidney cells growing in mice injected at the tumor site every 3 d with 100 μg of either the IgG1 control antibody (black box) or the IL6-neutralizing antibody (red box). (*) P < 0.01. (D) Representative tumors of RasG12V-transformed human kidney cells growing in mice injected every 3 d with 100 μg of either the IgG1 control antibody (control Ab) or the IL6-neutralizing antibody (αIL6 Ab).
Comment in
-
Targeting oncogenic Ras.Genes Dev. 2007 Aug 15;21(16):1989-92. doi: 10.1101/gad.1587907. Genes Dev. 2007. PMID: 17699748 Review. No abstract available.
References
-
- Aarden L.A., De Groot E.R., Schaap O.L., Lansdorp P.M., De Groot E.R., Schaap O.L., Lansdorp P.M., Schaap O.L., Lansdorp P.M., Lansdorp P.M. Production of hybridoma growth factor by human monocytes. Eur. J. Immunol. 1987;17:1411–1416. - PubMed
-
- Adams G.P., Weiner L.M., Weiner L.M. Monoclonal antibody therapy of cancer. Nat. Biotechnol. 2005;23:1147–1157. - PubMed
-
- Barber M.D., Fearon K.C., Ross J.A., Fearon K.C., Ross J.A., Ross J.A. Relationship of serum levels of interleukin-6, soluble interleukin-6 receptor and tumour necrosis factor receptors to the acute-phase protein response in advanced pancreatic cancer. Clin. Sci. 1999;96:83–87. - PubMed
-
- Cohen T., Nahari D., Cerem L.W., Neufeld G., Levi B.Z., Nahari D., Cerem L.W., Neufeld G., Levi B.Z., Cerem L.W., Neufeld G., Levi B.Z., Neufeld G., Levi B.Z., Levi B.Z. Interleukin 6 induces the expression of vascular endothelial growth factor. J. Biol. Chem. 1996;271:736–741. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials