

The essential peptidoglycan glycosyltransferase MurG forms a complex with proteins involved in lateral envelope growth as well as with proteins involved in cell division in Escherichia coli
- PMID: 17640276
- PMCID: PMC2170320
- DOI: 10.1111/j.1365-2958.2007.05851.x
The essential peptidoglycan glycosyltransferase MurG forms a complex with proteins involved in lateral envelope growth as well as with proteins involved in cell division in Escherichia coli
Abstract
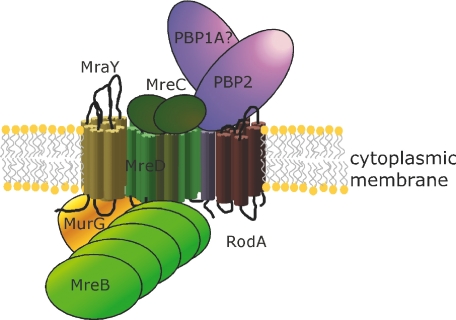
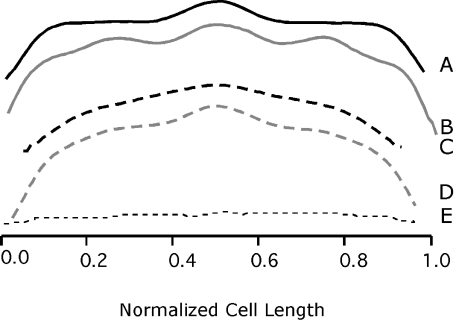
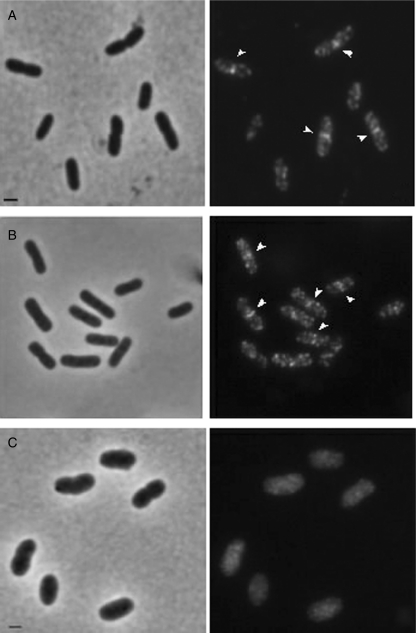
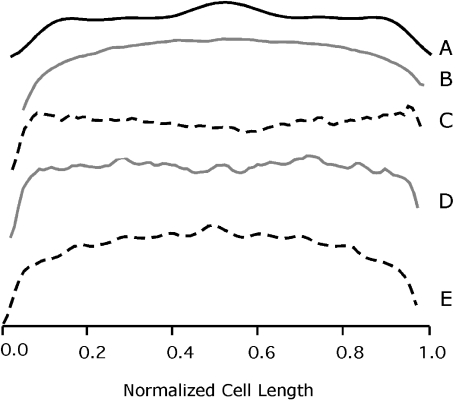
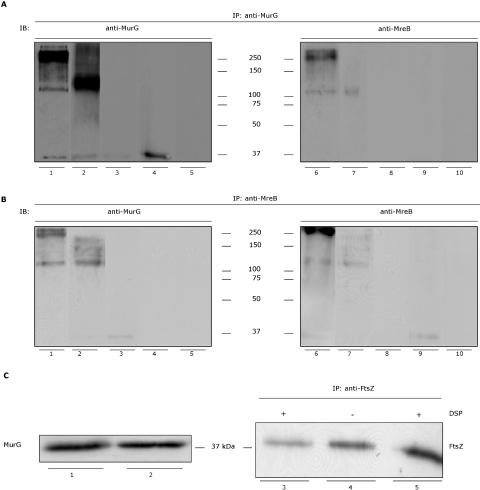
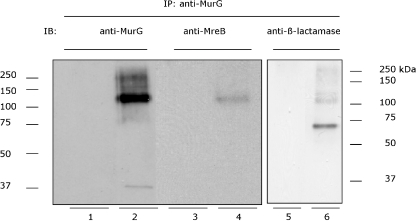
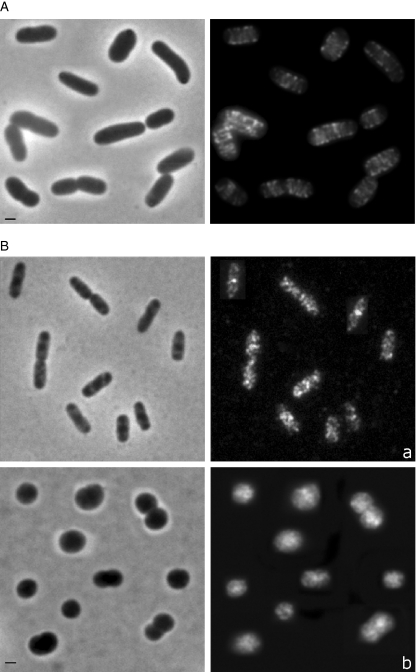
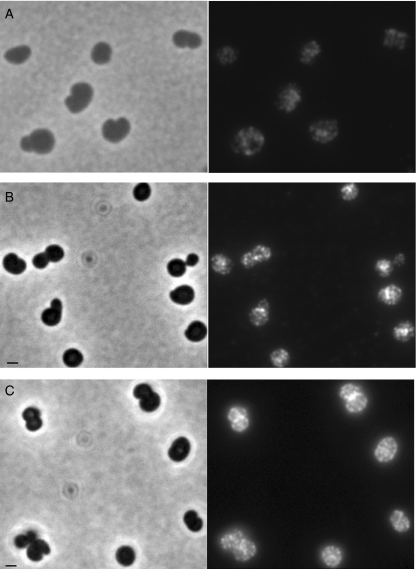
In Escherichia coli many enzymes including MurG are directly involved in the synthesis and assembly of peptidoglycan. MurG is an essential glycosyltransferase catalysing the last intracellular step of peptidoglycan synthesis. To elucidate its role during elongation and division events, localization of MurG using immunofluorescence microscopy was performed. MurG exhibited a random distribution in the cell envelope with a relatively higher intensity at the division site. This mid-cell localization was dependent on the presence of a mature divisome. Its localization in the lateral cell wall appeared to require the presence of MreCD. This could be indicative of a potential interaction between MurG and other proteins. Investigating this by immunoprecipitation revealed the association of MurG with MreB and MraY in the same protein complex. In view of this, the loss of rod shape of DeltamreBCD strain could be ascribed to the loss of MurG membrane localization. Consequently, this could prevent the localized supply of the lipid II precursor to the peptidoglycan synthesizing machinery involved in cell elongation. It is postulated that the involvement of MurG in the peptidoglycan synthesis concurs with two complexes, one implicated in cell elongation and the other in division. A model representing the first complex is proposed.
Figures
References
-
- Aaron M, Charbon G, Lam H, Schwarz H, Vollmer W, Jacobs-Wagner C. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Mol Microbiol. 2007;64:938–952. - PubMed
-
- Aarsman ME, Piette A, Fraipont C, Vinkenvleugel TM, Nguyen-Distèche M, den Blaauwen T. Maturation of the Escherichia coli divisome occurs in two steps. Mol Microbiol. 2005;55:1631–1645. - PubMed
-
- Addinall SG, Cao C, Lutkenhaus J. Temperature shift experiments with an FtsZ84(Ts) strain reveal rapid dynamics of FtsZ localization and indicate that the Z Ring is required throughout septation and cannot reoccupy division sites once constriction has initiated. J Bacteriol. 1997;179:4277–4284. - PMC - PubMed
-
- Auger G, van Heijenoort J, Mengin-Lecreulx D, Blanot D. A MurG assay which utilises a synthetic analogue of lipid I. FEMS Microbiol Lett. 2003;219:115–119. - PubMed
-
- Bertsche U, Kast T, Wolf B, Fraipont C, Aarsman ME, Kannenberg K, et al. Interaction between two murein (peptidoglycan) synthases, PBP3 and PBP1B, in Escherichia coli. Mol Microbiol. 2006;61:675–690. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
