

In situ dissection of a Polycomb response element in Drosophila melanogaster
- PMID: 17640916
- PMCID: PMC1941339
- DOI: 10.1073/pnas.0703144104
In situ dissection of a Polycomb response element in Drosophila melanogaster
Abstract
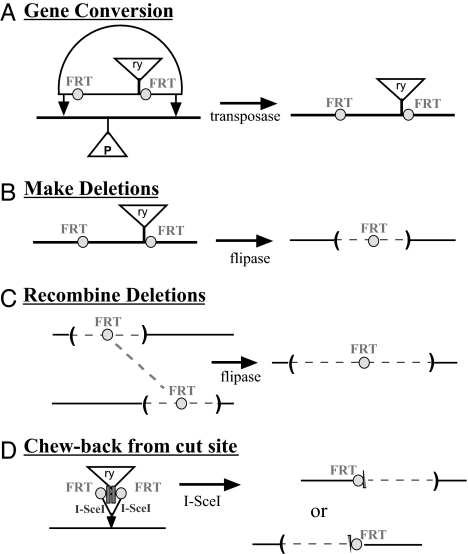
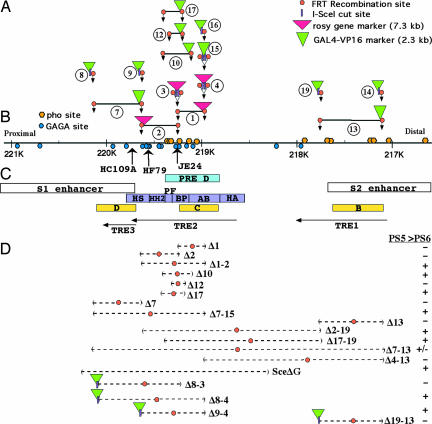
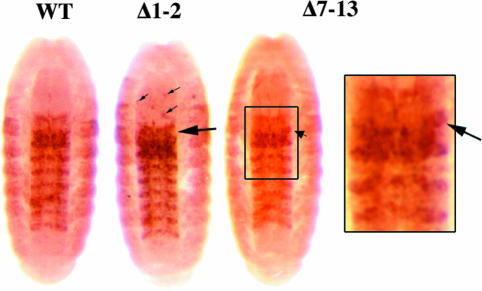
Genes of the Polycomb group maintain long-term, segment-specific repression of the homeotic genes in Drosophila. DNA targets of Polycomb group proteins, called Polycomb response elements (PREs), have been defined by several assays, but they have not been dissected in their original chromosomal context. An enhanced method of gene conversion was developed to generate a series of small, targeted deletions encompassing the best-studied PRE, upstream of the Ultrabithorax (Ubx) transcription unit in the bithorax complex. Deletions that removed an essential 185-bp core of the PRE caused anterior misexpression of Ubx and posterior segmental transformations, including the conversion of the third thoracic segment toward a duplicate first abdominal segment. These phenotypes were variable, suggesting some cooperation between this PRE and others in the bithorax complex. Larger deletions up to 3 kb were also created, which removed DNA sites reportedly needed for Ubx activation, including putative trithorax response elements. These deletions resulted in neither loss of Ubx expression nor loss-of-function phenotypes. Thus, the 3-kb region including the PRE is required for repression, but not for activation, of Ubx.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
The role of Polycomb-group response elements in regulation of engrailed transcription in Drosophila.Development. 2008 Feb;135(4):669-76. doi: 10.1242/dev.014779. Epub 2008 Jan 16. Development. 2008. PMID: 18199580
-
Replacement of a Drosophila Polycomb response element core, and in situ analysis of its DNA motifs.Mol Genet Genomics. 2008 Jun;279(6):595-603. doi: 10.1007/s00438-008-0336-3. Epub 2008 Mar 19. Mol Genet Genomics. 2008. PMID: 18350319
-
Specific combinations of boundary element and Polycomb response element are required for the regulation of the Hox genes in Drosophila melanogaster.Mech Dev. 2015 Nov;138 Pt 2:141-150. doi: 10.1016/j.mod.2015.07.016. Epub 2015 Aug 5. Mech Dev. 2015. PMID: 26254901
-
FLC: a hidden polycomb response element shows up in silence.Plant Cell Physiol. 2012 May;53(5):785-93. doi: 10.1093/pcp/pcr163. Epub 2011 Nov 22. Plant Cell Physiol. 2012. PMID: 22107881 Review.
-
The quest for mammalian Polycomb response elements: are we there yet?Chromosoma. 2016 Jun;125(3):471-96. doi: 10.1007/s00412-015-0539-4. Epub 2015 Oct 9. Chromosoma. 2016. PMID: 26453572 Free PMC article. Review.
Cited by
-
The Drosophila melanogaster Mutants apblot and apXasta Affect an Essential apterous Wing Enhancer.G3 (Bethesda). 2015 Apr 2;5(6):1129-43. doi: 10.1534/g3.115.017707. G3 (Bethesda). 2015. PMID: 25840432 Free PMC article.
-
Different transcriptional responses by the CRISPRa system in distinct types of heterochromatin in Drosophila melanogaster.Sci Rep. 2022 Jul 9;12(1):11702. doi: 10.1038/s41598-022-15944-7. Sci Rep. 2022. PMID: 35810197 Free PMC article.
-
Transcription through Polycomb response elements does not induce a switch from repression to activation.Proc Natl Acad Sci U S A. 2015 Dec 1;112(48):14755-6. doi: 10.1073/pnas.1520102112. Epub 2015 Nov 13. Proc Natl Acad Sci U S A. 2015. PMID: 26567151 Free PMC article. No abstract available.
-
Associating cellular epigenetic models with human phenotypes.Nat Rev Genet. 2017 Jul;18(7):441-451. doi: 10.1038/nrg.2017.32. Epub 2017 May 30. Nat Rev Genet. 2017. PMID: 28555657 Review.
-
Formation of a Polycomb-Domain in the Absence of Strong Polycomb Response Elements.PLoS Genet. 2016 Jul 28;12(7):e1006200. doi: 10.1371/journal.pgen.1006200. eCollection 2016 Jul. PLoS Genet. 2016. PMID: 27466807 Free PMC article.
References
-
- Jürgens G. Nature. 1985;316:153–155.
-
- Simon J. Curr Opin Cell Biol. 1995;7:376–385. - PubMed
-
- Akasaka T, van Lohuizen M, van der Lugt N, Mizutqani-Koseki Y, Kanno M, Taniguchi M, Vidal M, Alkema M, Berns A, Koseki H. Development (Cambridge, UK) 2001;128:1587–1597. - PubMed
-
- Pirrotta V. Trends Genet. 1997;13:314–318. - PubMed
-
- Simon J, Chiang A, Bender W, Shimell MJ, O'Connor M. Dev Biol. 1993;158:131–144. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
