

Review
doi: 10.1113/jphysiol.2007.136200.
Epub 2007 Jul 19.
The continuing case for the Renshaw cell
Affiliations
- PMID: 17640932
- PMCID: PMC2277064
- DOI: 10.1113/jphysiol.2007.136200
Item in Clipboard
Review
The continuing case for the Renshaw cell
J Physiol.
.
Abstract
Renshaw cell properties have been studied extensively for over 50 years, making them a uniquely well-defined class of spinal interneuron. Recent work has revealed novel ways to identify Renshaw cells in situ and this in turn has promoted a range of studies that have determined their ontogeny and organization of synaptic inputs in unprecedented detail. In this review we illustrate how mature Renshaw cell properties and connectivity arise through a combination of activity-dependent and genetically specified mechanisms. These new insights should aid the development of experimental strategies to manipulate Renshaw cells in spinal circuits and clarify their role in modulating motor output.
Figures
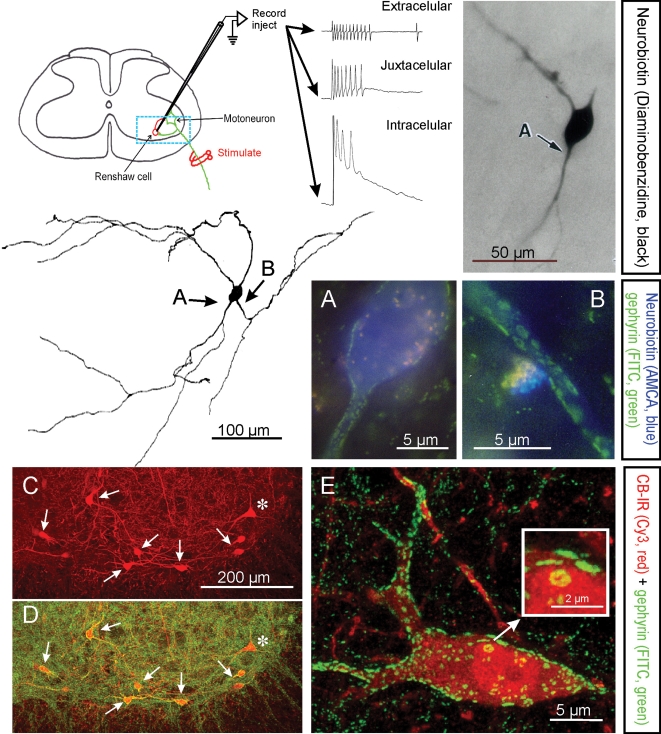
Renshaw cells are classically identified by their high-frequency discharge in response to antidromic motor axon action potentials recorded either extracellulary, juxtacellularly or intracellularly. The diagram shows the basic experimental set-up and resulting recordings. The typical high-frequency burst of action potentials recorded extracellularly is diminished following impalement of the cell but a long EPSP is revealed. The blue box indicates the approximate region of neuropil represented in C and D. Electrophysiologically identified Renshaw cells were injected with neurobiotin (Alvarez et al. 1997), which was initially revealed with α-methyl-coumarin (AMCA) to allow visualization of the pattern of gephyrin immunolabelling (FITC fluorochrome) on the recorded cell. Subsequent processing with diaminobenzidine permitted full reconstruction of the dendritic trees of the same cells. In this manner it was found that Renshaw cells are characterized by very large patches of gephyrin immunoreactivity in their proximal dendritic regions and somatic membrane; the frequency of these clusters drops markedly on more distal dendrites (Alvarez et al. 1997). A and B show at high magnification gephyrin-immunoreactive clusters in the regions indicated with arrows in the reconstructed neuron. C and D, Renshaw cells are also strongly immunoreactive (IR) for calbindin and the large majority of calbindin-IR cells in the ventral spinal cord display the large gephyrin clusters characteristic of Renshaw cells. In the confocal images shown in C and D only one calbindin-IR cell (*) did not display large gephyrin-IR clusters and was probably not a Renshaw cell. Non-Renshaw calbindin-IR cells tend to have larger cell bodies and do not receive strong inputs from cholinergic boutons. E, 2D projection of a 3D reconstructed calbindin-IR Renshaw cell displaying the characteristic large gephyrin clusters (note much larger clusters on the calbindin-IR cells than in the adjacent neuropil). One large cluster in the soma is shown magnified in the inset. Gephyrin-IR clusters represent inhibitory postsynaptic densities and the ones on Renshaw cells are among the larger found in the spinal cord.
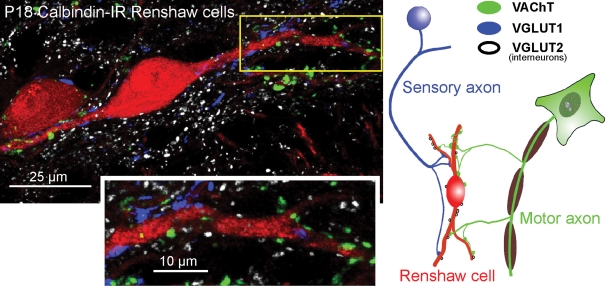
The confocal image (projection of 4 confocal optical planes; z-step = 0.5 μm) shows calbindin-IR (Cy3, red) Renshaw cells contacted by VAChT-IR (fluorescein-isothiocyanate (FITC), green), VGLUT1-IR (Alexa 405, blue) and VGLUT2-IR (Cy5, white) varicosities, summarized in the diagram at right. Individual confocal images were obtained using an Olympus FV1000 confocal microscope with excitation lines at 405, 488, 568 and 635 nm, and a 60 × oil (N.A. = 1.4) objective digitally zoomed 2 ×. Presumed contacts between varicosities and apposed dendrites are identified where varicosity immunofluorescence partially overlaps with dendrite calbindin immunofluorescence in a single optical plane. Most VGLUT1 and VAChT-IR inputs target dendrites, and the indicated, relatively proximal, dendritic segment (boxed area) is shown at higher magnification in the inset below. The density of VGLUT1 and VACHT-IR contacts increases distally (not shown; but see Fig. 3 for more distal contacts). Synaptic surface space on somata and most proximal dendrites is dominated by inhibitory synapses (see Fig. 1). VAChT and VGLUT1-IR varicosities are usually of relatively large size (see also Alvarez et al. 1999; Mentis et al. 2006), whereas VGLUT2-IR contacts tend to be of smaller size. Relative abundance of each type of excitatory input on Renshaw cells is on average VAChT > VGLUT2 > VGLUT1.
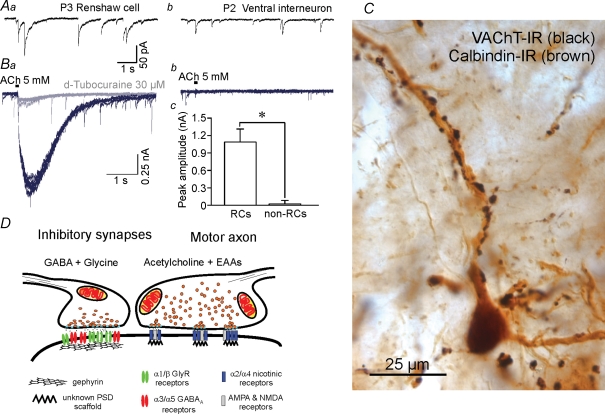
A, comparison of inhibitory currents in Renshaw cells and other spinal interneurons. Inhibitory mPSCs isolated pharmacologically with blockers of nicotinic, AMPA and NMDA receptors and recorded in the presence of tetradotoxin (see Gonzalez-Forero & Alvarez, 2005) display biphasic glycinergic (fast) and GABAergic (slow) components (Aa). Peak amplitudes are higher than in other interneurons (Ab) and dominated by the opening of glycine receptors. The decays have long GABAA-dependent components that correspond with high expression of α3/α5-containing GABAA receptors. Other ventral spinal interneurons generally lack these long GABAergic decays (Ab). B, Renshaw cells also display high postsynaptic sensitivity to acetylcholine (Ba and Bc) while other ventral interneurons are relatively insensitive (Bb and Bc) (D. Gonzalez-Forero and F. J. Alvarez, unpublished observations). Traces show responses to 30 ms pulses of 5 mm acetylcholine (ACh) from the same neurons from which records Aa and Ab were obtained before nicotinic receptor block. Recordings were performed on cells voltage-clamped at −75 mV in 0 Ca2+ and 0 Mg2+. All neurons with long GABAAR mPSCs exhibited ACh-evoked postsynaptic responses, whereas those with faster or no GABAergic events showed either small-amplitude or no responses (Bc). Postsynaptic responsiveness to ACh in Renshaw cells was effectively abolished with 30 μm d-tubocurarine (grey trace). C, motor axons imunolabelled with diamunobenzidine(DAB)–silver intensified VAChT immunoreactivity (black in the image) establish a large number of synapses around adult calbindin-IR Renshaw cell dendrites (brown in the image). Ultrastructurally each bouton displays multiple small active zones (F.J. Alvarez unpublished observations, not shown) and co-release acetylcholine and EAAs on PSDs that presumably co-localize nicotinic, AMPA and NMDA receptors (Mentis et al. 2005). These characteristics result in quite secure synapses with long EPSPs. D, the specific structural characteristics of each input on Renshaw cells correlate with their physiological properties. Inhibitory presynaptic boutons frequently co-release glycine and GABA and are opposite to large gephyrin clusters that accumulate large numbers of glycine and GABAA receptors in relatively large PSDs. It was concluded that the presence of strong and long-lasting excitatory motor axon inputs on Renshaw cells modulate the development of strong and long-lasting inhibitory synapses on these cells; the origin(s) of these inhibitory synapses have not yet been characterized (Geiman et al. 2000; Gonzalez-Forero et al. 2005).
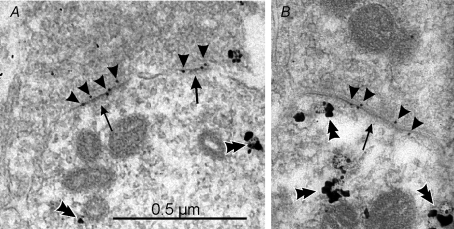
The images show calbindin-IR dendrites sampled from the ventral Renshaw cell region, immunostained in pre-embedding with immuno-nanogold and amplified with silver (double arrowheads). A proportion of postsynaptic densities (arrows) at excitatory synapses can be labelled with post-embedding immunogold (10 nm, arrowheads) for GluR4 AMPA receptor subunits in A and B. Similar results were obtained with antibodies against GluR2/3 or specific for GluR2. Although the combined pre- and postembedding immunocytochemistry and cryosubstitution techniques necessary to reveal these receptor subunits results in slightly compromised ultrastructure, particularly vesicle size and shape, the PSDs are asymmetric and contain AMPA receptors; hence, they are highly likely to represent excitatory (Type 1) synapses. Antibodies against GluR1 did not immunolabel any synapses on Renshaw cells, but intensely immunolabelled synapses in the dorsal horn. The images were obtained from a P20 animal.
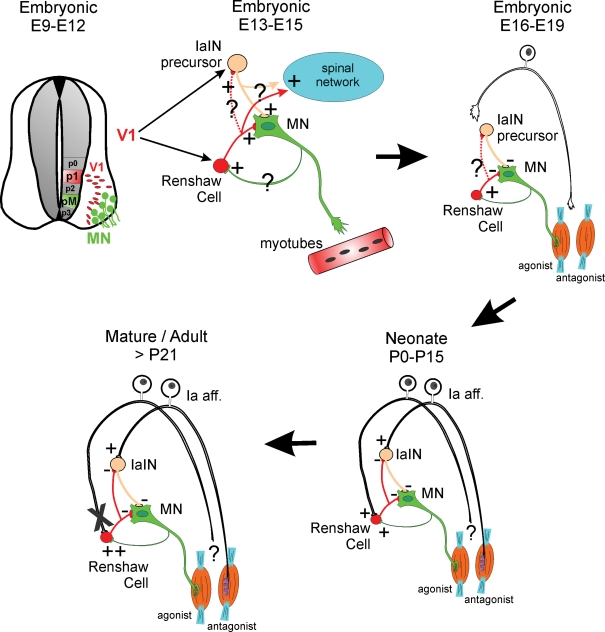
Renshaw cells are derived from V1 interneurons that are generated from p1 progenitors in the early neural tube (top, left). V1 interneurons take a ventral–lateral migratory path and position themselves in close relationship to motoneurons with whom they start to interact synaptically. V1 interneurons give rise to several classes of ipsilaterally projecting GABAergic/glycinergic premotor interneurons that include Renshaw cells and IaIN precursors (top, middle). At mid-embryonic stages all early connectivity (primarily cholinergic and GABAergic and less glycinergic and glutamatergic) is excitatory (Hanson & Landmesser, 2003; Myers et al. 2005); however, morphological details of these early connections are largely unknown (question marks). It is believed that this early connectivity has important roles in transmitting motoneuron spontaneous activity to the whole spinal network and that this spontaneous activity is important during early axonal guidance and target recognition. An amplifier role for early motoneuron-initiated spontaneous activity could be accomplished by Renshaw cells directly or indirectly through connections with other interneurons like IaINs. The anatomical nature and organization of these connections remains speculative at present. Current studies are actively pursuing clarification of this early synaptology. At late embryonic stages (top, right), inhibitory synapses become hyperpolarizing and the Renshaw cell's role in transmitting excitation down-regulates in favour of exerting inhibitory influences over the motoneurons. Primary afferent axons commence to invade the ventral horn of the spinal cord in late embryos at the same time that they induce the differentiation of the sensory apparatus in the periphery (muscle spindles and Golgi tendon organs). By the time of birth (bottom, right) Renshaw cells are contacted by both sensory afferents and motor axons and in this sense they are a class of proprioceptive inhibitory interneuron that also has direct excitation from motoneurons. The source of the sensory input is as yet unknown. If it originates from antagonist muscles, then Renshaw cells could behave at this stage as a special class of IaIN. After maturation and onset of weight-bearing locomotion (bottom, left) this sensory input is functionally lost (indicated by an X), although the sensory inputs are still present structurally, and the Renshaw cell's role then becomes more focused to recurrent inhibition, as it is in the adult.
References
-
- Alvarez FJ, Dewey DE, Harrington DA, Fyffe RE. Cell-type specific organization of glycine receptor clusters in the mammalian spinal cord. J Comp Neurol. 1997;379:150–170. - PubMed
-
- Alvarez FJ, Gonzalez-Forero D. Maturation of GABAA synapses on Renshaw cells. Abstr Soc Neurosci. 2006:716–717.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources