

Complete suppression of viral gene expression is associated with the onset and progression of lymphoid malignancy: observations in Bovine Leukemia Virus-infected sheep
- PMID: 17645797
- PMCID: PMC1948017
- DOI: 10.1186/1742-4690-4-51
Complete suppression of viral gene expression is associated with the onset and progression of lymphoid malignancy: observations in Bovine Leukemia Virus-infected sheep
Abstract
Background: During malignant progression, tumor cells need to acquire novel characteristics that lead to uncontrolled growth and reduced immunogenicity. In the Bovine Leukemia Virus-induced ovine leukemia model, silencing of viral gene expression has been proposed as a mechanism leading to immune evasion. However, whether proviral expression in tumors is completely suppressed in vivo was not conclusively demonstrated. Therefore, we studied viral expression in two selected experimentally-infected sheep, the virus or the disease of which had features that made it possible to distinguish tumor cells from their nontransformed counterparts.
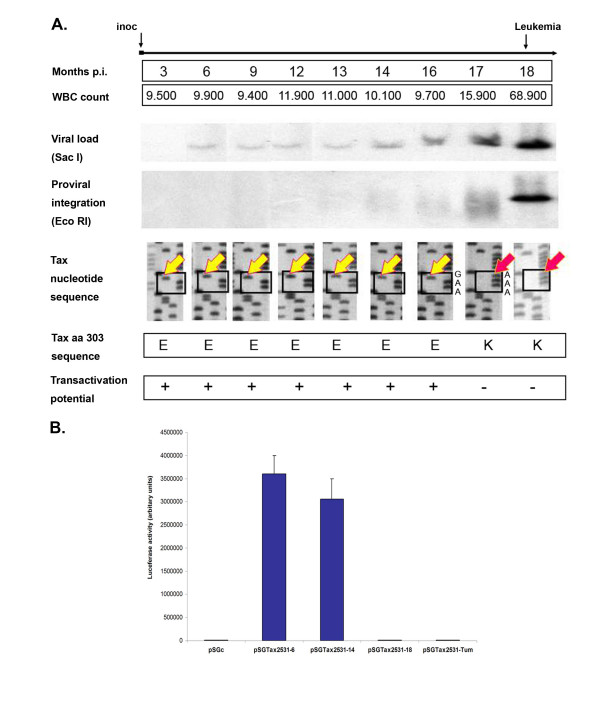
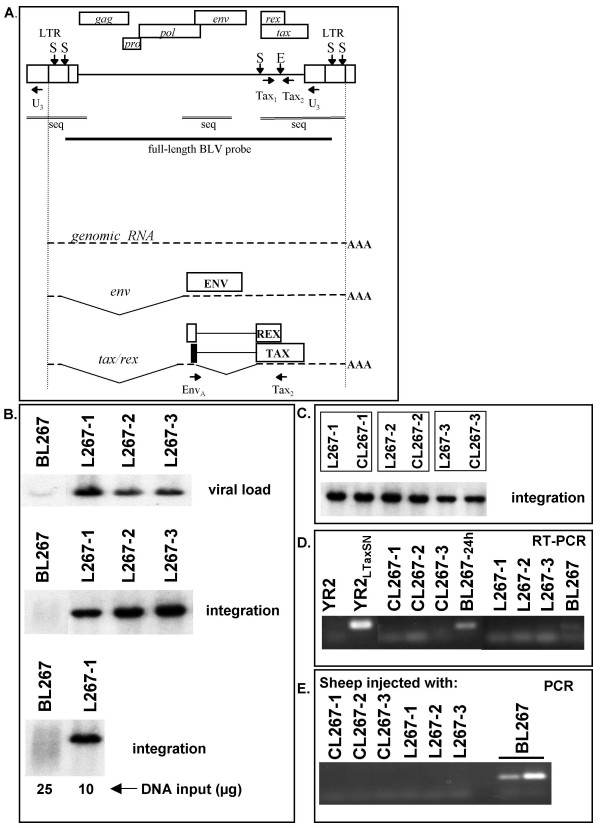
Results: In the first animal, we observed the emergence of a genetically modified provirus simultaneously with leukemia onset. We found a Tax-mutated (TaxK303) replication-deficient provirus in the malignant B-cell clone while functional provirus (TaxE303) had been consistently monitored over the 17-month aleukemic period. In the second case, both non-transformed and transformed BLV-infected cells were present at the same time, but at distinct sites. While there was potentially-active provirus in the non-leukemic blood B-cell population, as demonstrated by ex-vivo culture and injection into naïve sheep, virus expression was completely suppressed in the malignant B-cells isolated from the lymphoid tumors despite the absence of genetic alterations in the proviral genome. These observations suggest that silencing of viral genes, including the oncoprotein Tax, is associated with tumor onset.
Conclusion: Our findings suggest that silencing is critical for tumor progression and identify two distinct mechanisms-genetic and epigenetic-involved in the complete suppression of virus and Tax expression. We demonstrate that, in contrast to systems that require sustained oncogene expression, the major viral transforming protein Tax can be turned-off without reversing the transformed phenotype. We propose that suppression of viral gene expression is a contributory factor in the impairment of immune surveillance and the uncontrolled proliferation of the BLV-infected tumor cell.
Figures
References
-
- Marincola FM, Jaffee EM, Hicklin DJ, Ferrone S. Escape of human solid tumors from T-cell recognition: molecular mechanisms and functional significance. Adv Immunol. 2000;74:181–273. - PubMed
-
- Burny A, Willems,L.,Callebaut,I.,Adam,E.,Cludts,I, Dequiedt,F.,Droogmans,L.,Grimonpont,C.,Kerkhofs,P.,Mammerickx,M.,Portetelle,D.,Van den Broeke,A.,and Kettman,R. Bovine Leukemia Virus: biology and mode of transformation. Viruses and Cancer Minson, A C , Neil, J C and McRae, M A (eds), Cambridge University Press, Cambridge. 1994. pp. 313–334.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
