

The role of the Runx transcription factors in thymocyte differentiation and in homeostasis of naive T cells
- PMID: 17646406
- PMCID: PMC2118679
- DOI: 10.1084/jem.20070133
The role of the Runx transcription factors in thymocyte differentiation and in homeostasis of naive T cells
Erratum in
- J Exp Med. 2008 Aug 4;205(8):1939
Abstract
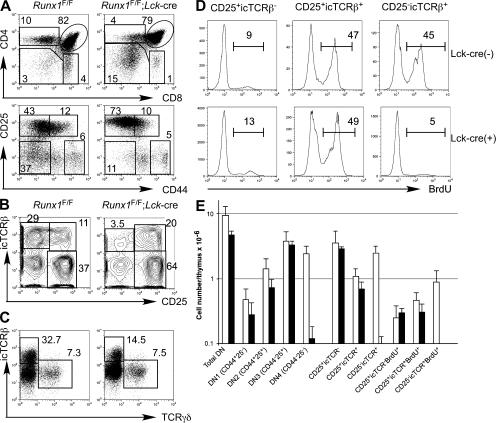
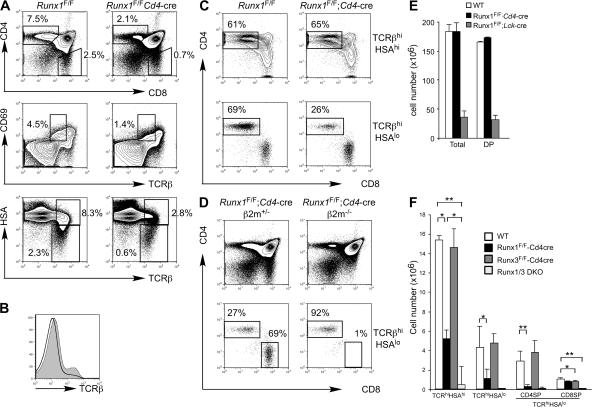
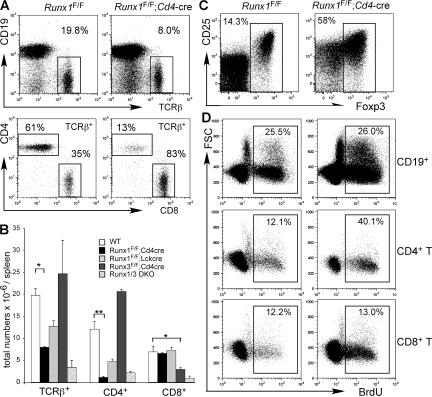
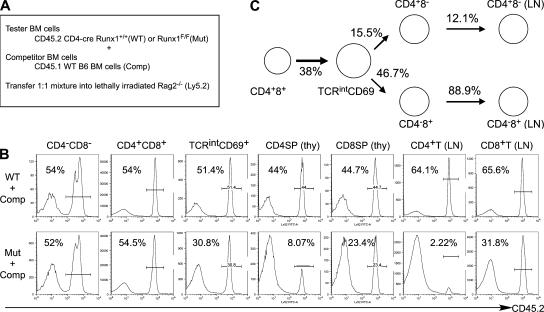
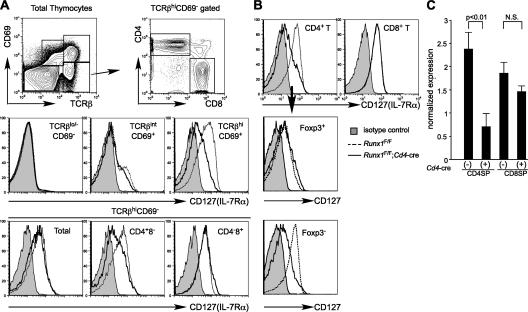
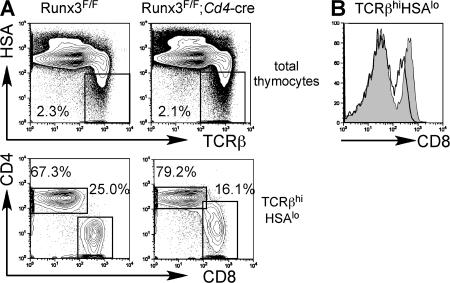
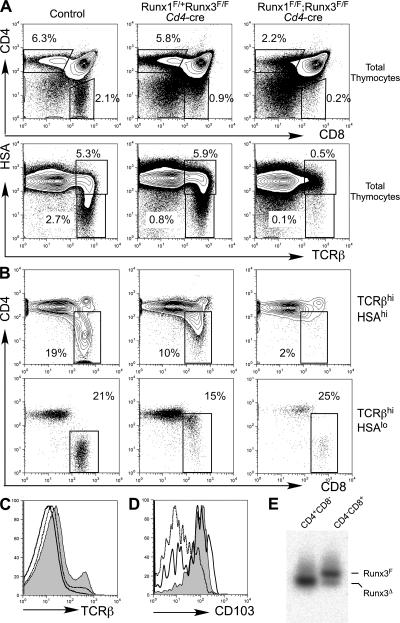
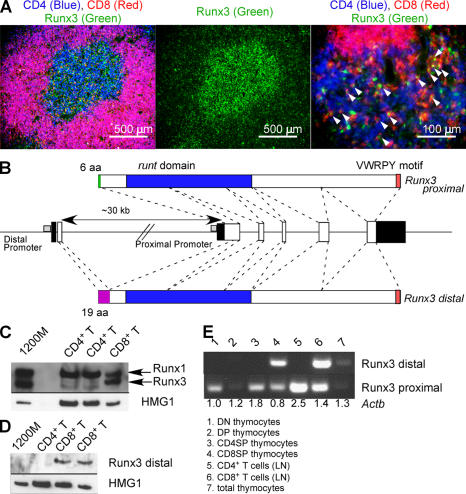
Members of the Runx family of transcriptional regulators are required for the appropriate expression of CD4 and CD8 at discrete stages of T cell development. The roles of these factors in other aspects of T cell development are unknown. We used a strategy to conditionally inactivate the genes encoding Runx1 or Runx3 at different stages of thymocyte development, demonstrating that Runx1 regulates the transitions of developing thymocytes from the CD4(-)CD8(-) double-negative stage to the CD4(+)CD8(+) double-positive (DP) stage and from the DP stage to the mature single-positive stage. Runx1 and Runx3 deficiencies caused marked reductions in mature thymocytes and T cells of the CD4(+) helper and CD8(+) cytotoxic T cell lineages, respectively. Runx1-deficient CD4(+) T cells had markedly reduced expression of the interleukin 7 receptor and exhibited shorter survival. In addition, inactivation of both Runx1 and Runx3 at the DP stages resulted in a severe block in development of CD8(+) mature thymocytes. These results indicate that Runx proteins have important roles at multiple stages of T cell development and in the homeostasis of mature T cells.
Figures
References
-
- Weerkamp, F., K. Pike-Overzet, and F.J. Staal. 2006. T-sing progenitors to commit. Trends Immunol. 27:125–131. - PubMed
-
- von Boehmer, H., I. Aifantis, F. Gounari, O. Azogui, L. Haughn, I. Apostolou, E. Jaeckel, F. Grassi, and L. Klein. 2003. Thymic selection revisited: how essential is it? Immunol. Rev. 191:62–78. - PubMed
-
- Godfrey, D.I., and A. Zlotnik. 1993. Control points in early T-cell development. Immunol. Today. 14:547–553. - PubMed
-
- Aifantis, I., M. Mandal, K. Sawai, A. Ferrando, and T. Vilimas. 2006. Regulation of T-cell progenitor survival and cell-cycle entry by the pre-T-cell receptor. Immunol. Rev. 209:159–169. - PubMed
-
- Hoffman, E.S., L. Passoni, T. Crompton, T.M. Leu, D.G. Schatz, A. Koff, M.J. Owen, and A.C. Hayday. 1996. Productive T-cell receptor beta-chain gene rearrangement: coincident regulation of cell cycle and clonality during development in vivo. Genes Dev. 10:948–962. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
