

Sexually dimorphic gene expression in the heart of mice and men
- PMID: 17646949
- PMCID: PMC2755745
- DOI: 10.1007/s00109-007-0240-z
Sexually dimorphic gene expression in the heart of mice and men
Abstract
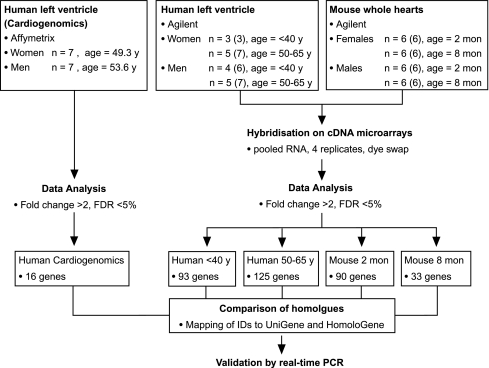
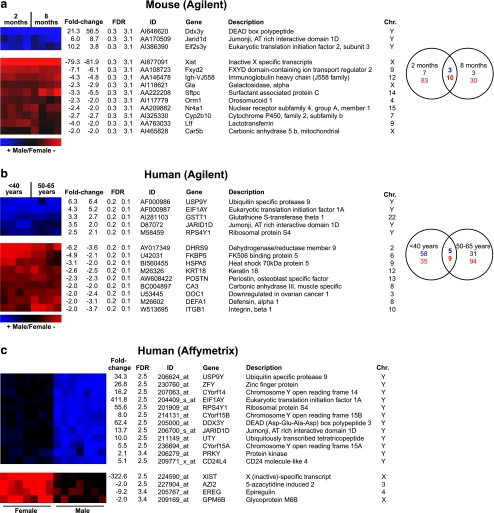
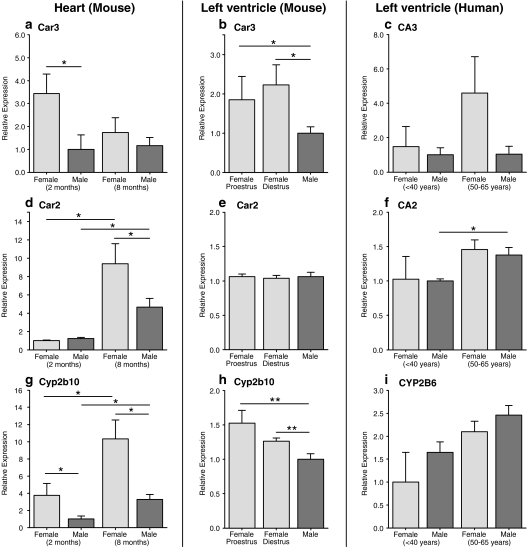
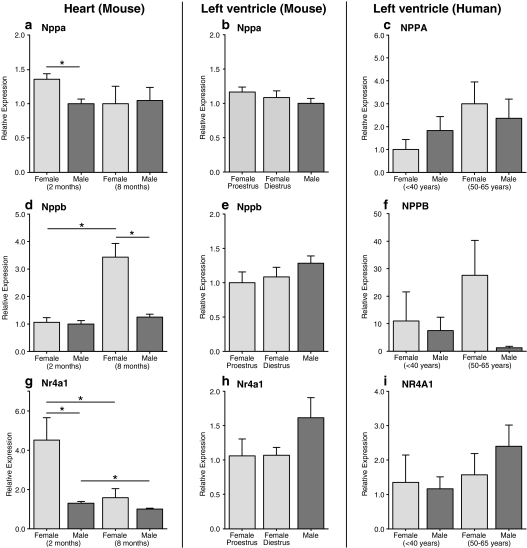
The prevalence and clinical manifestation of several cardiovascular diseases vary considerably with sex and age. Thus, a better understanding of the molecular basis of these differences may represent a starting point for an improved gender-specific medicine. Despite the fact that sex-specific differences have been observed in the cardiovascular system of humans and animal models, systematic analyses of sexual dimorphisms at the transcriptional level in the healthy heart are missing. Therefore we performed gene expression profiling on mouse and human cardiac samples of both sexes and young as well as aged individuals and verified our results for a subset of genes using real-time polymerase chain reaction in independent left ventricular samples. To tackle the question whether sex differences are evolutionarily conserved, we also compared sexually dimorphic genes between both species. We found that genes located on sex chromosomes were the most abundant ones among the sexually dimorphic genes. Male-specific expression of Y-linked genes was observed in mouse hearts as well as in the human myocardium (e.g. Ddx3y, Eif2s3y and Jarid1d). Higher expression levels of X-linked genes were detected in female mice for Xist, Timp1 and Car5b and XIST, EIF2S3X and GPM6B in women. Furthermore, genes on autosomal chromosomes encoding cytochromes of the monoxygenase family (e.g. Cyp2b10), carbonic anhydrases (e.g. Car2 and Car3) and natriuretic peptides (e.g. Nppb) were identified with sex- and/or age-specific expression levels. This study underlines the relevance of sex and age as modifiers of cardiac gene expression.
Figures
References
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/j.devcel.2004.05.005', 'is_inner': False, 'url': 'https://doi.org/10.1016/j.devcel.2004.05.005'}, {'type': 'PubMed', 'value': '15177028', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/15177028/'}]}
- Rinn JL, Rozowsky JS, Laurenzi IJ, Petersen PH, Zou K, Zhong W, Gerstein M, Snyder M (2004) Major molecular differences between mammalian sexes are involved in drug metabolism and renal function. Dev Cell 6:791–800 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1210/me.2003-0138', 'is_inner': False, 'url': 'https://doi.org/10.1210/me.2003-0138'}, {'type': 'PubMed', 'value': '14684848', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/14684848/'}]}
- Ahluwalia A, Clodfelter KH, Waxman DJ (2004) Sexual dimorphism of rat liver gene expression: regulatory role of growth hormone revealed by deoxyribonucleic acid microarray analysis. Mol Endocrinol 18:747–760 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1101/gr.5217506', 'is_inner': False, 'url': 'https://doi.org/10.1101/gr.5217506'}, {'type': 'PMC', 'value': 'PMC1524872', 'is_inner': False, 'url': 'https://pmc.ncbi.nlm.nih.gov/articles/PMC1524872/'}, {'type': 'PubMed', 'value': '16825664', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/16825664/'}]}
- Yang X, Schadt EE, Wang S, Wang H, Arnold AP, Ingram-Drake L, Drake TA, Lusis AJ (2006) Tissue-specific expression and regulation of sexually dimorphic genes in mice. Genome Res 16:995–1004 - PMC - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/j.tig.2005.03.005', 'is_inner': False, 'url': 'https://doi.org/10.1016/j.tig.2005.03.005'}, {'type': 'PubMed', 'value': '15851067', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/15851067/'}]}
- Rinn JL, Snyder M (2005) Sexual dimorphism in mammalian gene expression. Trends Genet 21:298–305 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/S0008-6363(00)00005-5', 'is_inner': False, 'url': 'https://doi.org/10.1016/s0008-6363(00)00005-5'}, {'type': 'PubMed', 'value': '10727651', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/10727651/'}]}
- Hayward CS, Kelly RP, Collins P (2000) The roles of gender, the menopause and hormone replacement on cardiovascular function. Cardiovasc Res 46:28–49 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
