

Regulation of stress-induced intracellular sorting and chaperone function of Hsp27 (HspB1) in mammalian cells
- PMID: 17650072
- PMCID: PMC2275061
- DOI: 10.1042/BJ20070195
Regulation of stress-induced intracellular sorting and chaperone function of Hsp27 (HspB1) in mammalian cells
Abstract
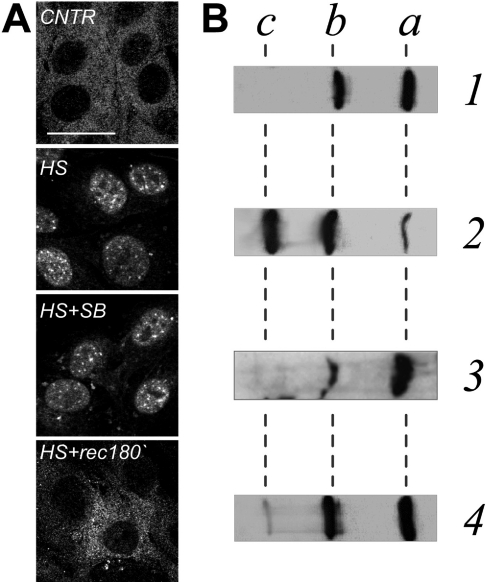
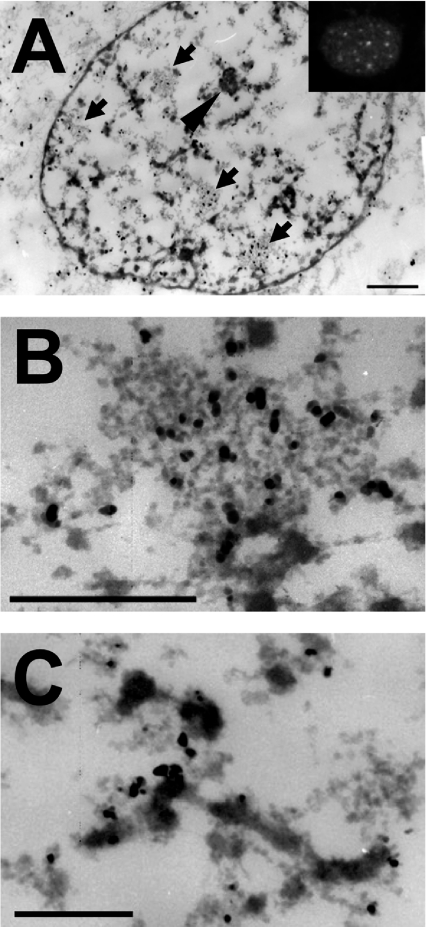
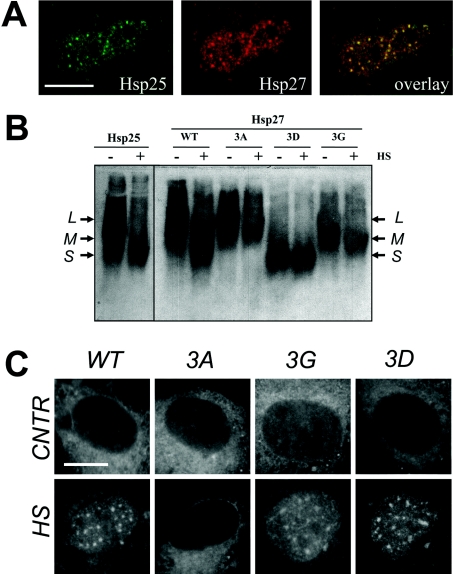
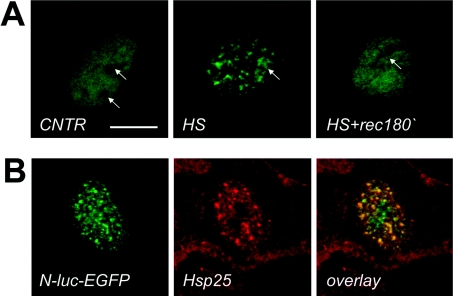
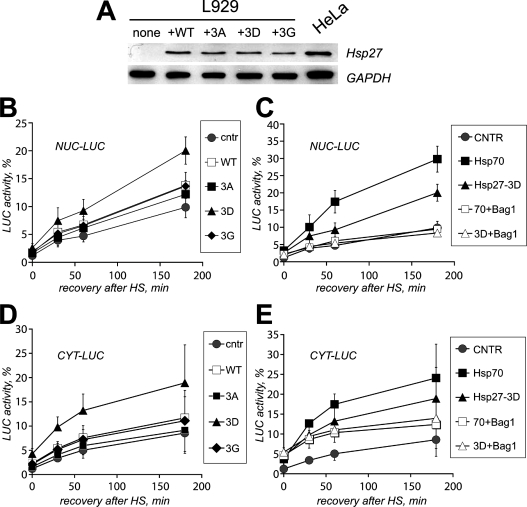
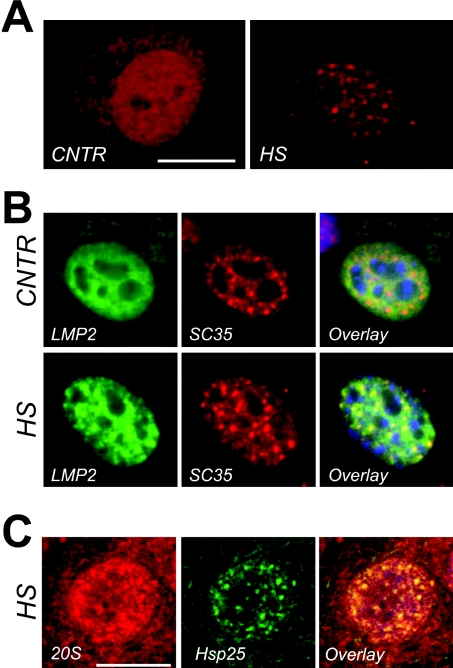
In vitro, small Hsps (heat-shock proteins) have been shown to have chaperone function capable of keeping unfolded proteins in a form competent for Hsp70-dependent refolding. However, this has never been confirmed in living mammalian cells. In the present study, we show that Hsp27 (HspB1) translocates into the nucleus upon heat shock, where it forms granules that co-localize with IGCs (interchromatin granule clusters). Although heat-induced changes in the oligomerization status of Hsp27 correlate with its phosphorylation and nuclear translocation, Hsp27 phosphorylation alone is not sufficient for effective nuclear translocation of HspB1. Using firefly luciferase as a heat-sensitive reporter protein, we demonstrate that HspB1 expression in HspB1-deficient fibroblasts enhances protein refolding after heat shock. The positive effect of HspB1 on refolding is completely diminished by overexpression of Bag-1 (Bcl-2-associated athanogene), the negative regulator of Hsp70, consistent with the idea of HspB1 being the substrate holder for Hsp70. Although HspB1 and luciferase both accumulate in nuclear granules after heat shock, our results suggest that this is not related to the refolding activity of HspB1. Rather, granular accumulation may reflect a situation of failed refolding where the substrate is stored for subsequent degradation. Consistently, we found 20S proteasomes concentrated in nuclear granules of HspB1 after heat shock. We conclude that HspB1 contributes to an increased chaperone capacity of cells by binding unfolded proteins that are hereby kept competent for refolding by Hsp70 or that are sorted to nuclear granules if such refolding fails.
Figures
References
-
- Lavoie J. N., Gingras-Breton G., Tanguay R. M., Landry J. Induction of Chinese hamster HSP27 gene expression in mouse cells confers resistance to heat shock. HSP27 stabilization of the microfilament organization. J. Biol. Chem. 1993;268:3420–3429. - PubMed
-
- Jakob U., Gaestel M., Engel K., Buchner J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993;268:1517–1520. - PubMed
-
- Ehrnsperger M., Buchner J., Gaestel M. Structure and function of small heat-shock proteins. In: Fink A., Goto Y., editors. Molecular chaperones in the life cycle of proteins. New York: Marcel Dekker, Inc.; 1997. pp. 533–575.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous