

Improving water use in crop production
- PMID: 17652070
- PMCID: PMC2610175
- DOI: 10.1098/rstb.2007.2175
Improving water use in crop production
Abstract
Globally, agriculture accounts for 80-90% of all freshwater used by humans, and most of that is in crop production. In many areas, this water use is unsustainable; water supplies are also under pressure from other users and are being affected by climate change. Much effort is being made to reduce water use by crops and produce 'more crop per drop'. This paper examines water use by crops, taking particularly a physiological viewpoint, examining the underlying relationships between carbon uptake, growth and water loss. Key examples of recent progress in both assessing and improving crop water productivity are described. It is clear that improvements in both agronomic and physiological understanding have led to recent increases in water productivity in some crops. We believe that there is substantial potential for further improvements owing to the progress in understanding the physiological responses of plants to water supply, and there is considerable promise within the latest molecular genetic approaches, if linked to the appropriate environmental physiology. We conclude that the interactions between plant and environment require a team approach looking across the disciplines from genes to plants to crops in their particular environments to deliver improved water productivity and contribute to sustainability.
Figures
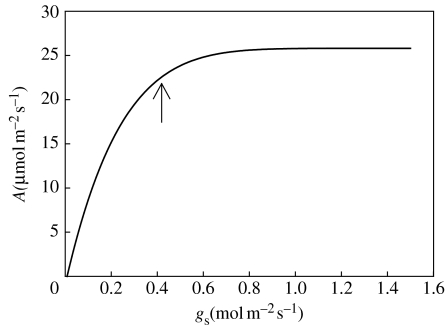
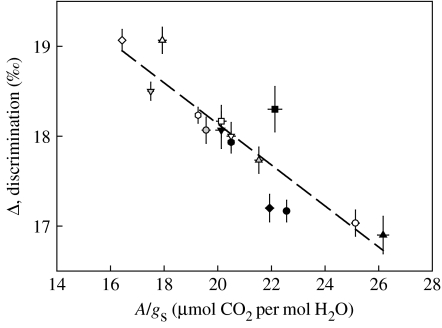

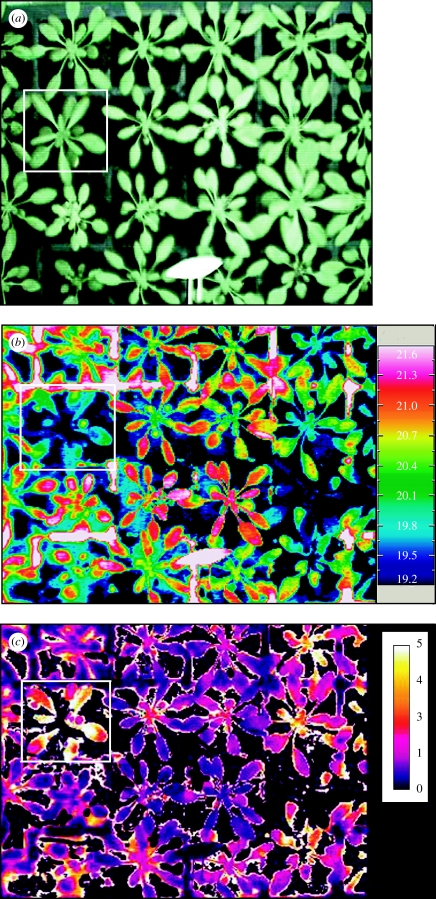
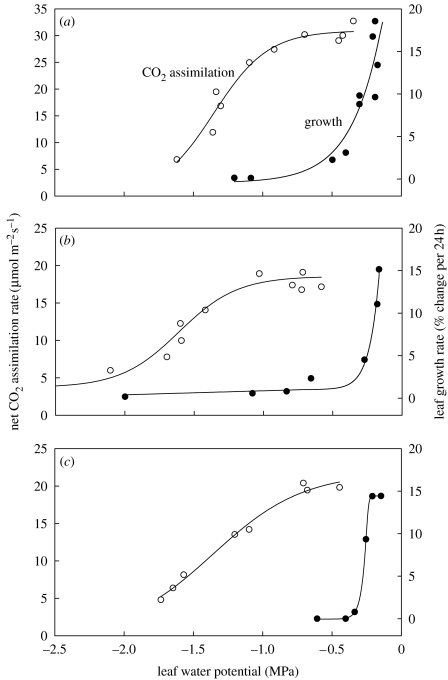
References
-
- Alonso-Blanco C, Koornneef M. Naturally occurring variation in Arabidopsis: an underexploited resource for plant genetics. Trends Plant Sci. 2000;5:22–29. doi:10.1016/S1360-1385(99)01510-1 - DOI - PubMed
-
- Araus J.L. The problems of sustainable water use in the Mediterranean and research requirements agriculture. Ann. Appl. Biol. 2004;144:259–272. doi:10.1111/j.1744-7348.2004.tb00342.x - DOI
-
- Bacon M.A. Water use efficiency in plant biology. Blackwell Publishing; Oxford, UK: 2004. p. 327.
-
- Baker N.R, Oxborough K. Chlorophyll fluorescence as a probe of photosynthetic productivity. In: Papageorgiou G.C, Govindjee , editors. Chlorophyll a fluorescence: a signature of photosynthesis. Springer; Dordrecht, The Netherelands: 2004. pp. 65–82.
-
- Baker N.R, Rosenqvist E. Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J. Exp. Bot. 2004;55:1607–1621. doi:10.1093/jxb/erh196 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
