

The planar polarity pathway promotes coordinated cell migration during Drosophila oogenesis
- PMID: 17652348
- PMCID: PMC1991286
- DOI: 10.1242/dev.010447
The planar polarity pathway promotes coordinated cell migration during Drosophila oogenesis
Abstract
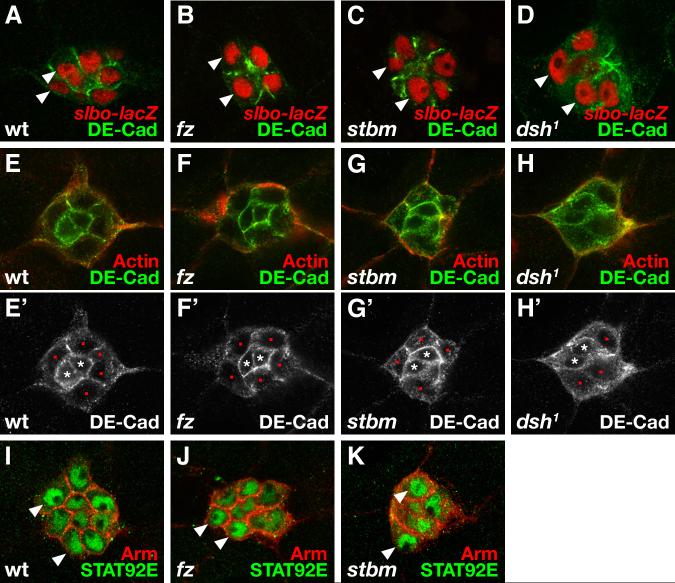
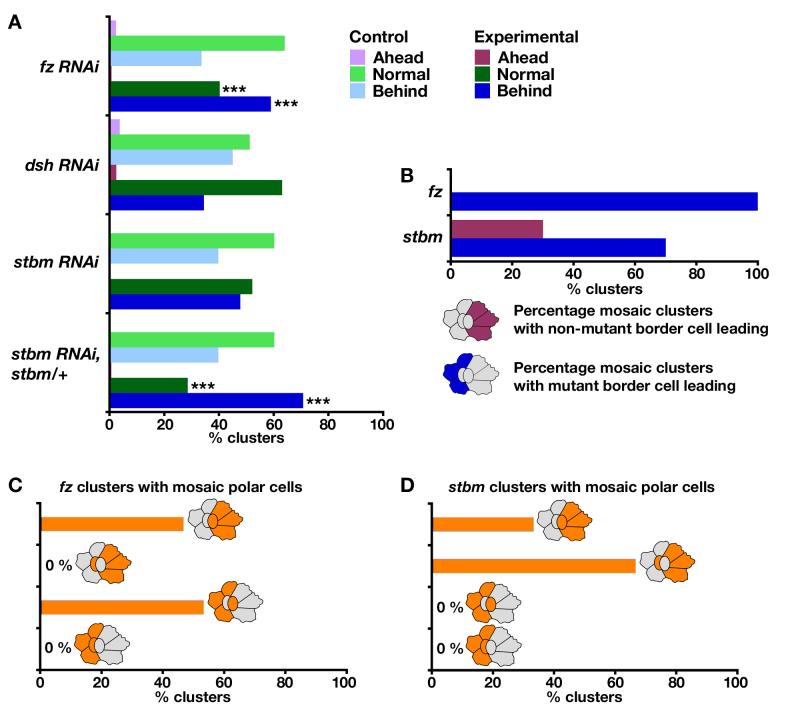
Cell migration is fundamental in both animal morphogenesis and disease. The migration of individual cells is relatively well-studied; however, in vivo, cells often remain joined by cell-cell junctions and migrate in cohesive groups. How such groups of cells coordinate their migration is poorly understood. The planar polarity pathway coordinates the polarity of non-migrating cells in epithelial sheets and is required for cell rearrangements during vertebrate morphogenesis. It is therefore a good candidate to play a role in the collective migration of groups of cells. Drosophila border cell migration is a well-characterised and genetically tractable model of collective cell migration, during which a group of about six to ten epithelial cells detaches from the anterior end of the developing egg chamber and migrates invasively towards the oocyte. We find that the planar polarity pathway promotes this invasive migration, acting both in the migrating cells themselves and in the non-migratory polar follicle cells that they carry along. Disruption of planar polarity signalling causes abnormalities in actin-rich processes on the cell surface and leads to less-efficient migration. This is apparently due, in part, to a loss of regulation of Rho GTPase activity by the planar polarity receptor Frizzled, which itself becomes localised to the migratory edge of the border cells. We conclude that, during collective cell migration, the planar polarity pathway can mediate communication between motile and non-motile cells, which enhances the efficiency of migration via the modulation of actin dynamics.
Figures




References
-
- Adler PN, Zhu C, Stone D. Inturned localizes to the proximal side of wing cells under the instruction of upstream planar polarity proteins. Curr. Biol. 2004;14:2046–2051. - PubMed
-
- Bastock R, Strutt H, Strutt D. Strabismus is asymmetrically localised and binds to Prickle and Dishevelled during Drosophila planar polarity patterning. Development. 2003;130:3007–3014. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Chen X, Oh SW, Zheng Z, Chen HW, Shin HH, Hou SX. Cyclin D-Cdk4 and cyclin E-Cdk2 regulate the Jak/STAT signal transduction pathway in Drosophila. Dev. Cell. 2003;4:179–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
