

Analysis of the VSG gene silent archive in Trypanosoma brucei reveals that mosaic gene expression is prominent in antigenic variation and is favored by archive substructure
- PMID: 17652423
- PMCID: PMC1950903
- DOI: 10.1101/gr.6421207
Analysis of the VSG gene silent archive in Trypanosoma brucei reveals that mosaic gene expression is prominent in antigenic variation and is favored by archive substructure
Abstract
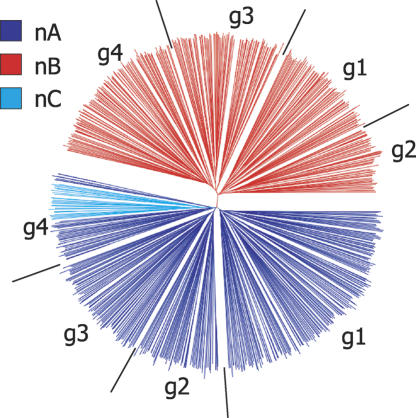
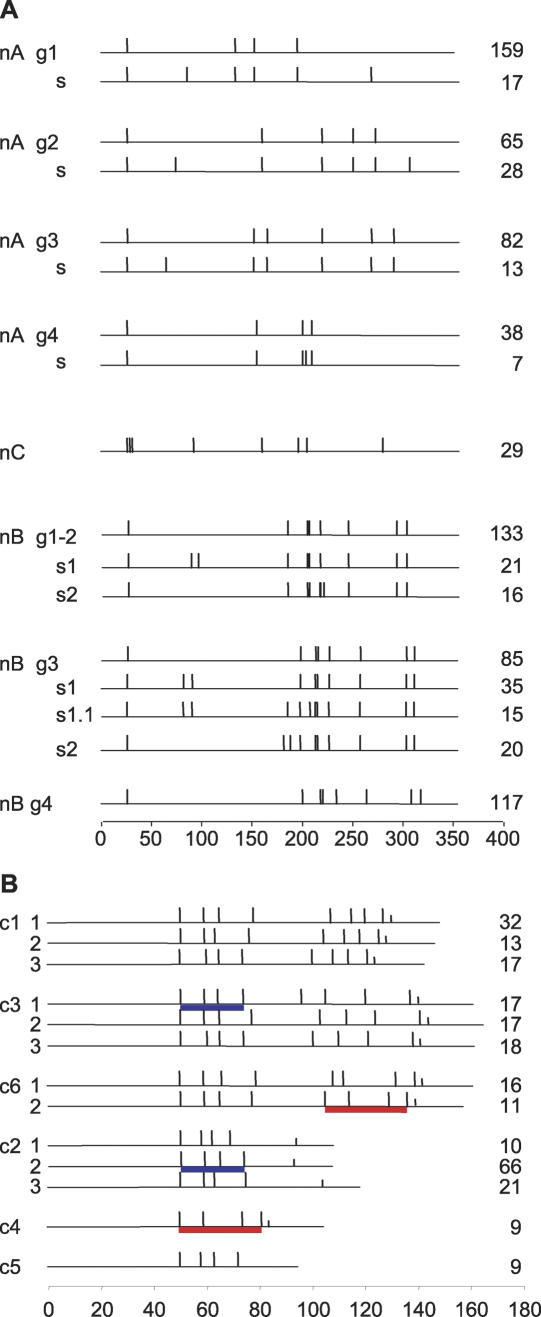
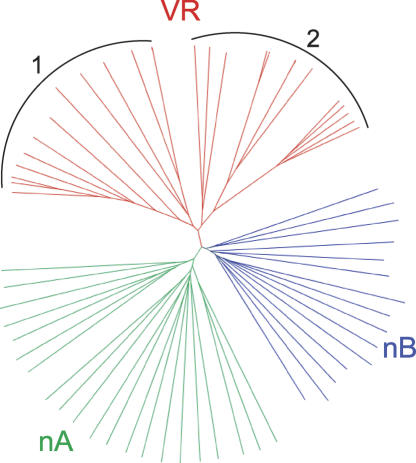
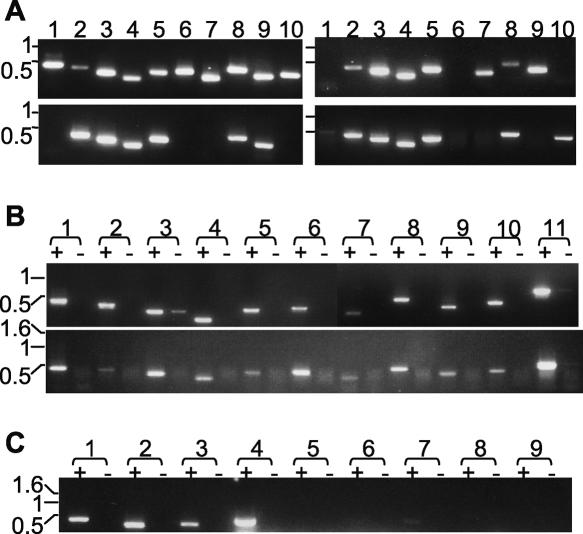
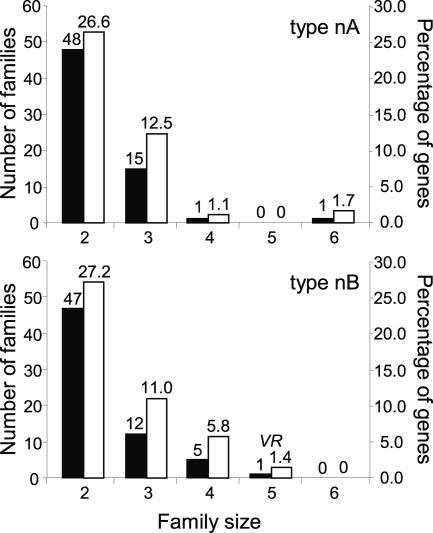
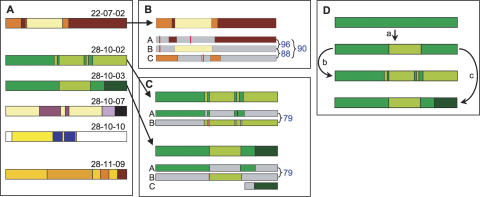
Trypanosoma brucei evades host acquired immunity through differential activation of its large archive of silent variant surface glycoprotein (VSG) genes, most of which are pseudogenes in subtelomeric arrays. We have analyzed 940 VSGs, representing one half to two thirds of the arrays. Sequence types A and B of the VSG N-terminal domains were confirmed, while type C was found to be a constituent of type A. Two new C-terminal domain types were found. Nearly all combinations of domain types occurred, with some bias to particular combinations. One-third of encoded N-terminal domains, but only 13% of C-terminal domains, are intact, indicating a particular need for silent VSGs to gain a functional C-terminal domain to be expressed. About 60% of VSGs are unique, the rest occurring in subfamilies of two to four close homologs (>50%-52% peptide identity). We found a subset of VSG-related genes, differing from VSGs in genomic environment and expression patterns, and predict they have distinct function. Almost all (92%) full-length array VSGs have the partially conserved flanks associated with the duplication mechanism that activates silent genes, and these sequences have also contributed to archive evolution, mediating most of the conversions of segments, containing >/=1 VSG, within and between arrays. During infection, intact array genes became activated by duplication after two weeks, and mosaic VSGs assembled from pseudogenes became expressed by week three and predominated by week four. The small subfamily structure of the archive appears to be fundamental in providing the interacting donors for mosaic formation.
Figures

References
-
- Aline R., Macdonald G., Brown E., Allison J., Myler P., Rothwell V., Stuart K., Macdonald G., Brown E., Allison J., Myler P., Rothwell V., Stuart K., Brown E., Allison J., Myler P., Rothwell V., Stuart K., Allison J., Myler P., Rothwell V., Stuart K., Myler P., Rothwell V., Stuart K., Rothwell V., Stuart K., Stuart K. (TAA)n within sequences flanking several intrachromosomal variant surface glycoprotein genes in Trypanosoma brucei. Nucleic Acids Res. 1985;13:3161–3177. - PMC - PubMed
-
- Aury J.M., Jaillon O., Duret L., Noel B., Jubin C., Porcel B.M., Segurens B., Daubin V., Anthouard V., Aiach N., Jaillon O., Duret L., Noel B., Jubin C., Porcel B.M., Segurens B., Daubin V., Anthouard V., Aiach N., Duret L., Noel B., Jubin C., Porcel B.M., Segurens B., Daubin V., Anthouard V., Aiach N., Noel B., Jubin C., Porcel B.M., Segurens B., Daubin V., Anthouard V., Aiach N., Jubin C., Porcel B.M., Segurens B., Daubin V., Anthouard V., Aiach N., Porcel B.M., Segurens B., Daubin V., Anthouard V., Aiach N., Segurens B., Daubin V., Anthouard V., Aiach N., Daubin V., Anthouard V., Aiach N., Anthouard V., Aiach N., Aiach N., et al. Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia. Nature. 2006;444:171–178. - PubMed
-
- Barbet A.F., Kamper S.M., Kamper S.M. The importance of mosaic genes to trypanosome survival. Parasitol. Today. 1993;9:63–66. - PubMed
-
- Barry J.D., McCulloch R., McCulloch R. Antigenic variation in trypanosomes: Enhanced phenotypic variation in a eukaryotic parasite. Adv. Parasitol. 2001;49:1–70. - PubMed
-
- Barry J.D., Ginger M.L., Burton P., McCulloch R., Ginger M.L., Burton P., McCulloch R., Burton P., McCulloch R., McCulloch R. Why are parasite contingency genes often associated with telomeres? Int. J. Parasitol. 2003;33:29–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources