

tRNase Z catalysis and conserved residues on the carboxy side of the His cluster
- PMID: 17655328
- PMCID: PMC2526284
- DOI: 10.1021/bi700578v
tRNase Z catalysis and conserved residues on the carboxy side of the His cluster
Abstract
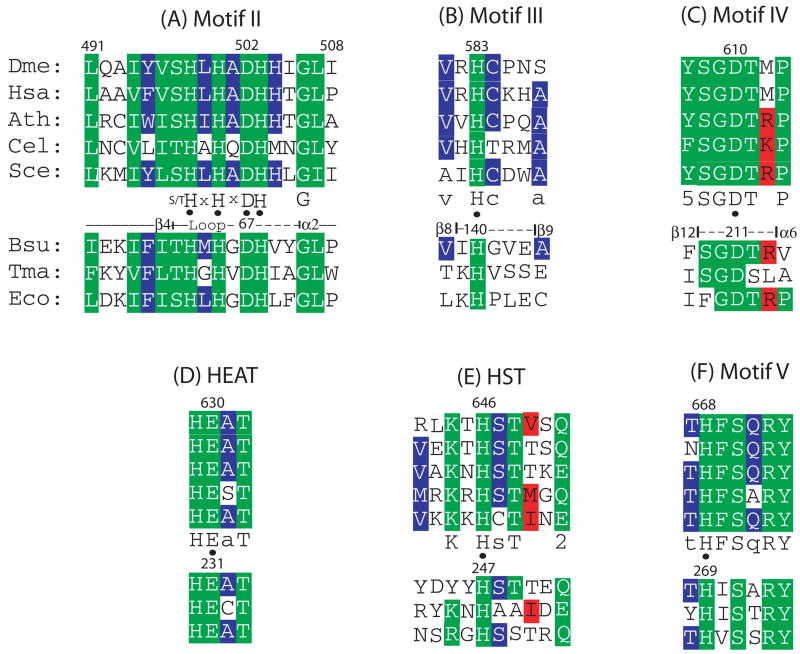
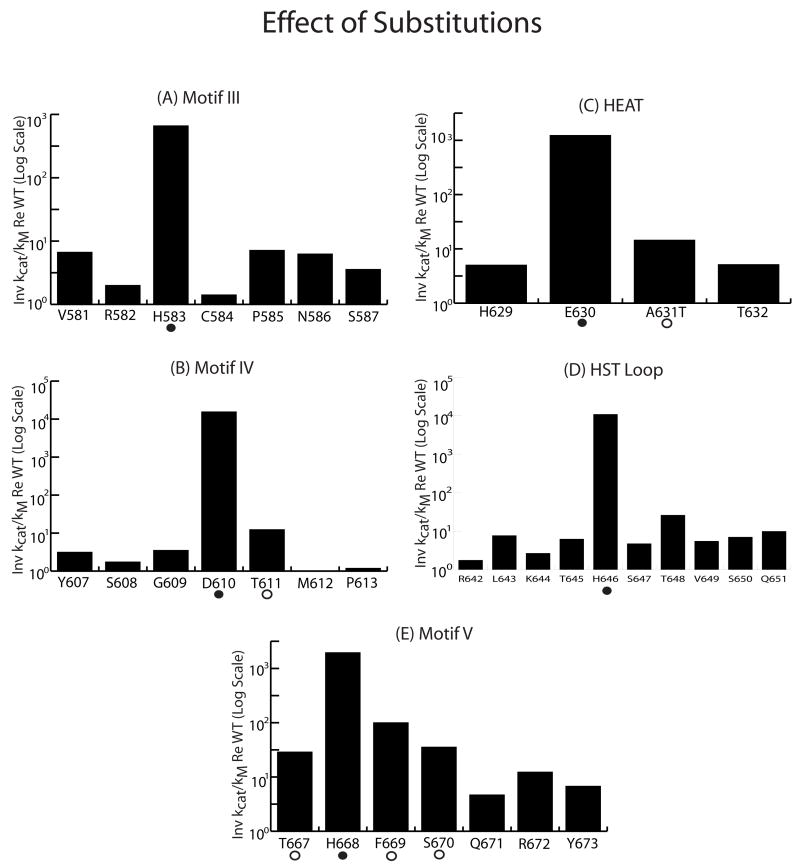
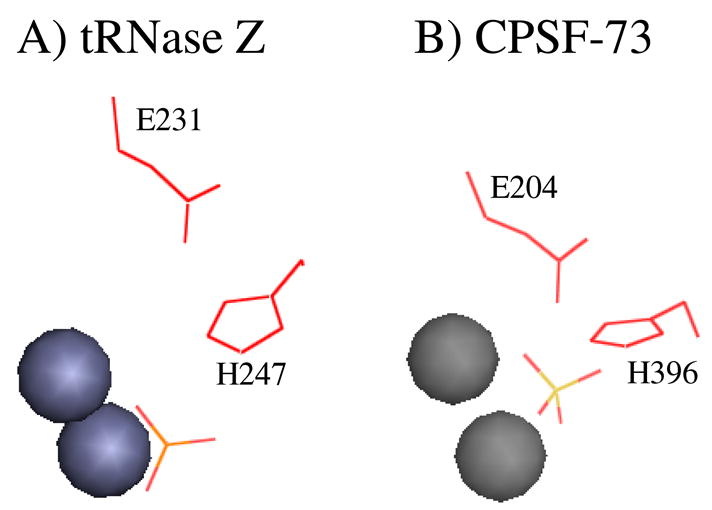
tRNAs are transcribed as precursors and processed in a series of required reactions leading to aminoacylation and translation. The 3'-end trailer can be removed by the pre-tRNA processing endonuclease tRNase Z, an ancient, conserved member of the beta-lactamase superfamily of metal-dependent hydrolases. The signature sequence of this family, the His domain (HxHxDH, Motif II), and histidines in Motifs III and V and aspartate in Motif IV contribute seven side chains for the coordination of two divalent metal ions. We previously investigated the effects on catalysis of substitutions in Motif II and in the PxKxRN loop and Motif I on the amino side of Motif II. Herein, we present the effects of substitutions on the carboxy side of Motif II within Motifs III, IV, the HEAT and HST loops, and Motif V. Substitution of the Motif IV aspartate reduces catalytic efficiency more than 10,000-fold. Histidines in Motif III, V, and the HST loop are also functionally important. Strikingly, replacement of Glu in the HEAT loop with Ala reduces efficiency by approximately 1000-fold. Proximity and orientation of this Glu side chain relative to His in the HST loop and the importance of both residues for catalysis suggest that they function as a duo in proton transfer at the final stage of reaction, characteristic of the tRNase Z class of RNA endonucleases.
Figures


Similar articles
-
Residues in two homology blocks on the amino side of the tRNase Z His domain contribute unexpectedly to pre-tRNA 3' end processing.RNA. 2006 Jun;12(6):1104-15. doi: 10.1261/rna.4206. Epub 2006 Apr 17. RNA. 2006. PMID: 16618969 Free PMC article.
-
Residues in the conserved His domain of fruit fly tRNase Z that function in catalysis are not involved in substrate recognition or binding.J Mol Biol. 2005 Jul 8;350(2):189-99. doi: 10.1016/j.jmb.2005.04.073. J Mol Biol. 2005. PMID: 15935379
-
Effect of changes in the flexible arm on tRNase Z processing kinetics.J Biol Chem. 2009 Jun 5;284(23):15685-91. doi: 10.1074/jbc.M900745200. Epub 2009 Apr 7. J Biol Chem. 2009. PMID: 19351879 Free PMC article.
-
The making of tRNAs and more - RNase P and tRNase Z.Prog Mol Biol Transl Sci. 2009;85:319-68. doi: 10.1016/S0079-6603(08)00808-8. Prog Mol Biol Transl Sci. 2009. PMID: 19215776 Review.
-
The tRNase Z family of proteins: physiological functions, substrate specificity and structural properties.Biol Chem. 2005 Dec;386(12):1253-64. doi: 10.1515/BC.2005.142. Biol Chem. 2005. PMID: 16336119 Review.
Cited by
-
Expanding the repertoire of human tandem repeat RNA-binding proteins.PLoS One. 2023 Sep 20;18(9):e0290890. doi: 10.1371/journal.pone.0290890. eCollection 2023. PLoS One. 2023. PMID: 37729217 Free PMC article.
-
Structure of PhnP, a phosphodiesterase of the carbon-phosphorus lyase pathway for phosphonate degradation.J Biol Chem. 2009 Jun 19;284(25):17216-17226. doi: 10.1074/jbc.M808392200. Epub 2009 Apr 14. J Biol Chem. 2009. PMID: 19366688 Free PMC article.
-
Identification and sequence analysis of metazoan tRNA 3'-end processing enzymes tRNase Zs.PLoS One. 2012;7(9):e44264. doi: 10.1371/journal.pone.0044264. Epub 2012 Sep 4. PLoS One. 2012. PMID: 22962606 Free PMC article.
-
Tethered domains and flexible regions in tRNase Z(L), the long form of tRNase Z.PLoS One. 2013 Jul 17;8(7):e66942. doi: 10.1371/journal.pone.0066942. Print 2013. PLoS One. 2013. PMID: 23874404 Free PMC article.
-
Pathogenesis-related mutations in the T-loops of human mitochondrial tRNAs affect 3' end processing and tRNA structure.RNA Biol. 2012 Mar;9(3):283-91. doi: 10.4161/rna.19025. Epub 2012 Mar 1. RNA Biol. 2012. PMID: 22336717 Free PMC article.
References
-
- de la Sierra-Gallay IL, Pellegrini O, Condon C. Structural basis for substrate binding, cleavage and allostery in the tRNA maturase RNase Z. Nature. 2005;433:657–661. - PubMed
-
- de la Sierra-Gallay IL, Mathy N, Pellegrini O, Condon C. Structure of the ubiquitous 3′ processing enzyme RNase Z bound to transfer RNA. Nat Struct Mol Biol. 2006;13:376–377. - PubMed
-
- DeLano WL. The PyMOL Molecular Graphics System on World Wide Web. 2002. http://www.pymol.org.
-
- Dominski Z, Yang XC, Marzluff WF. The polyadenylation factor CPSF-73 is involved in histone-pre-mRNA processing. Cell. 2005;123:37–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
