

Telomerase and Tel1p preferentially associate with short telomeres in S. cerevisiae
- PMID: 17656141
- PMCID: PMC2650483
- DOI: 10.1016/j.molcel.2007.07.016
Telomerase and Tel1p preferentially associate with short telomeres in S. cerevisiae
Abstract
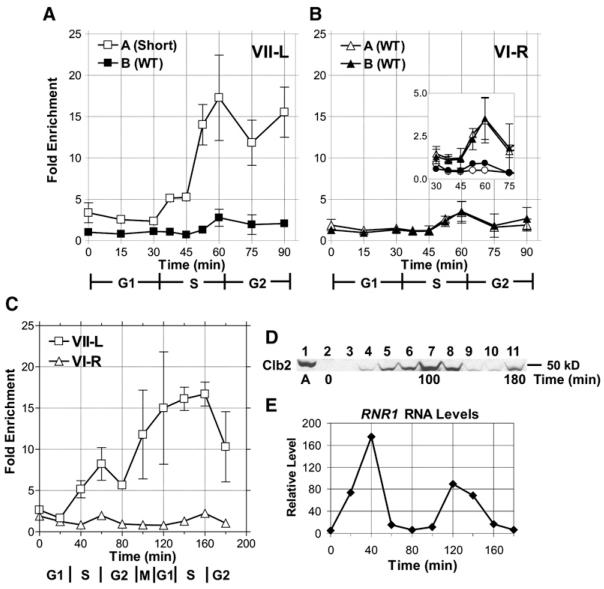
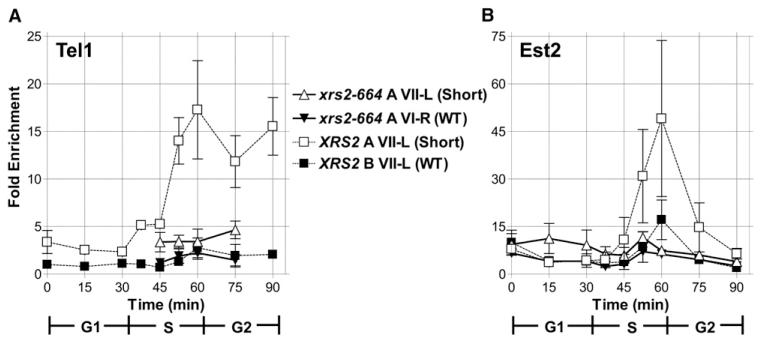
In diverse organisms, telomerase preferentially elongates short telomeres. We generated a single short telomere in otherwise wild-type (WT) S. cerevisiae cells. The binding of the positive regulators Ku and Cdc13p was similar at short and WT-length telomeres. The negative regulators Rif1p and Rif2p were present at the short telomere, although Rif2p levels were reduced. Two telomerase holoenzyme components, Est1p and Est2p, were preferentially enriched at short telomeres in late S/G2 phase, the time of telomerase action. Tel1p, the yeast ATM-like checkpoint kinase, was highly enriched at short telomeres from early S through G2 phase and even into the next cell cycle. Nonetheless, induction of a single short telomere did not elicit a cell-cycle arrest. Tel1p binding was dependent on Xrs2p and required for preferential binding of telomerase to short telomeres. These data suggest that Tel1p targets telomerase to the DNA ends most in need of extension.
Figures
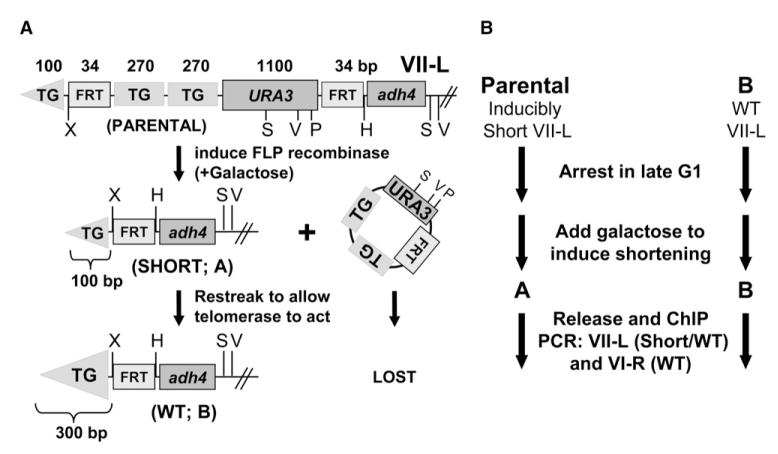
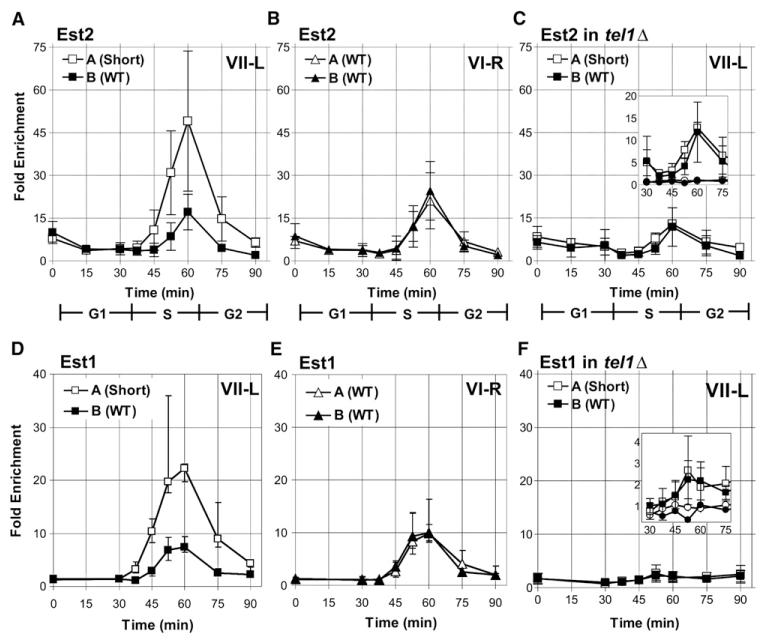
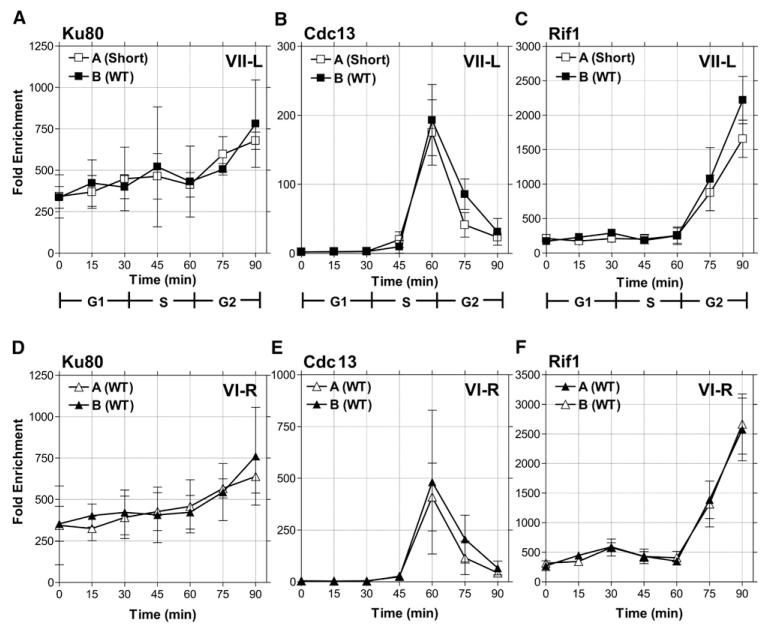
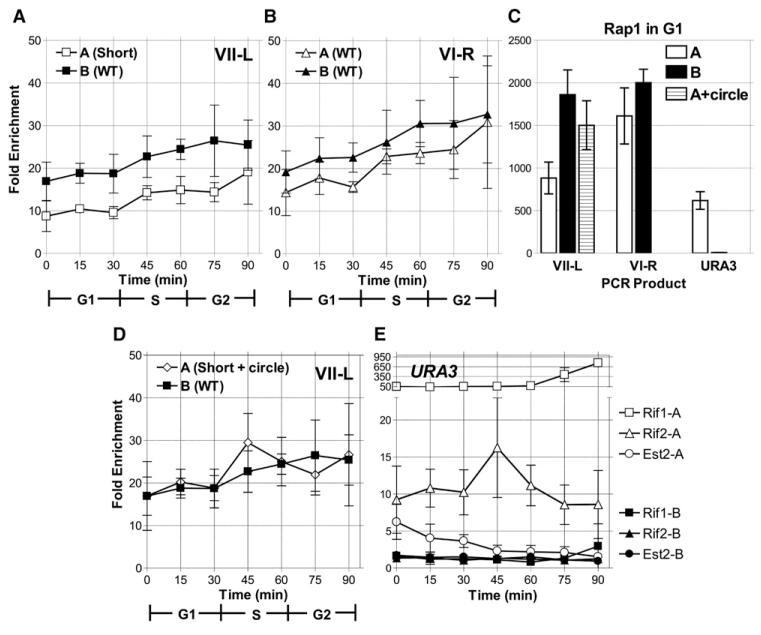
References
-
- Belli G, Gari E, Aldea M, Herrero E. Functional analysis of yeast essential genes using a promoter-substitution cassette and the tetracycline-regulatable dual expression system. Yeast. 1998;14:1127–1138. - PubMed
-
- Bianchi A, Negrini S, Shore D. Delivery of yeast telomerase to a DNA break depends on the recruitment functions of Cdc13 and Est1. Mol. Cell. 2004;16:139–146. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
